fetching data ... 
Background: Sjögren’s syndrome (SS) is a systemic autoimmune disease characterized by lymphocytic infiltration of exocrine glands and heterogeneous extraglandular manifestations. Cell-type–specific gene regulation may influence SS susceptibility, but causal immune-cell–resolved regulatory genes remain incompletely defined.
Objectives: To investigate the causal role of immune-cell–specific gene expression in SS risk.
Methods: Instrumental variables for Mendelian randomization (MR) were derived from conditionally independent cis-expression quantitative trait loci (cis-eQTLs) identified in the OneK1K single-cell eQTL (sc-eQTL) dataset [1]. Genome-wide significant cis-eQTLs (P < 5.0 × 10 −8 ) were LD-clumped using the 1000 Genomes European reference panel (r 2 < 0.001; 10 Mb). Exposure and outcome summary statistics were harmonized, excluding ambiguous or strand-incompatible variants; palindromic variants were retained only when allele frequencies allowed unambiguous alignment. Weak instruments (F < 10) were removed. Two-sample MR was performed to estimate the causal effects of immune-cell–specific gene expression on SS risk using large-scale GWAS summary statistics (FinnGen R12). Bayesian colocalization was used to assess whether cis-eQTL and SS association signals shared a causal variant, prioritizing associations with strong evidence of colocalization (PP.H4 > 80%). Effect estimates are reported as odds ratios (ORs) with 95% confidence intervals (CIs).
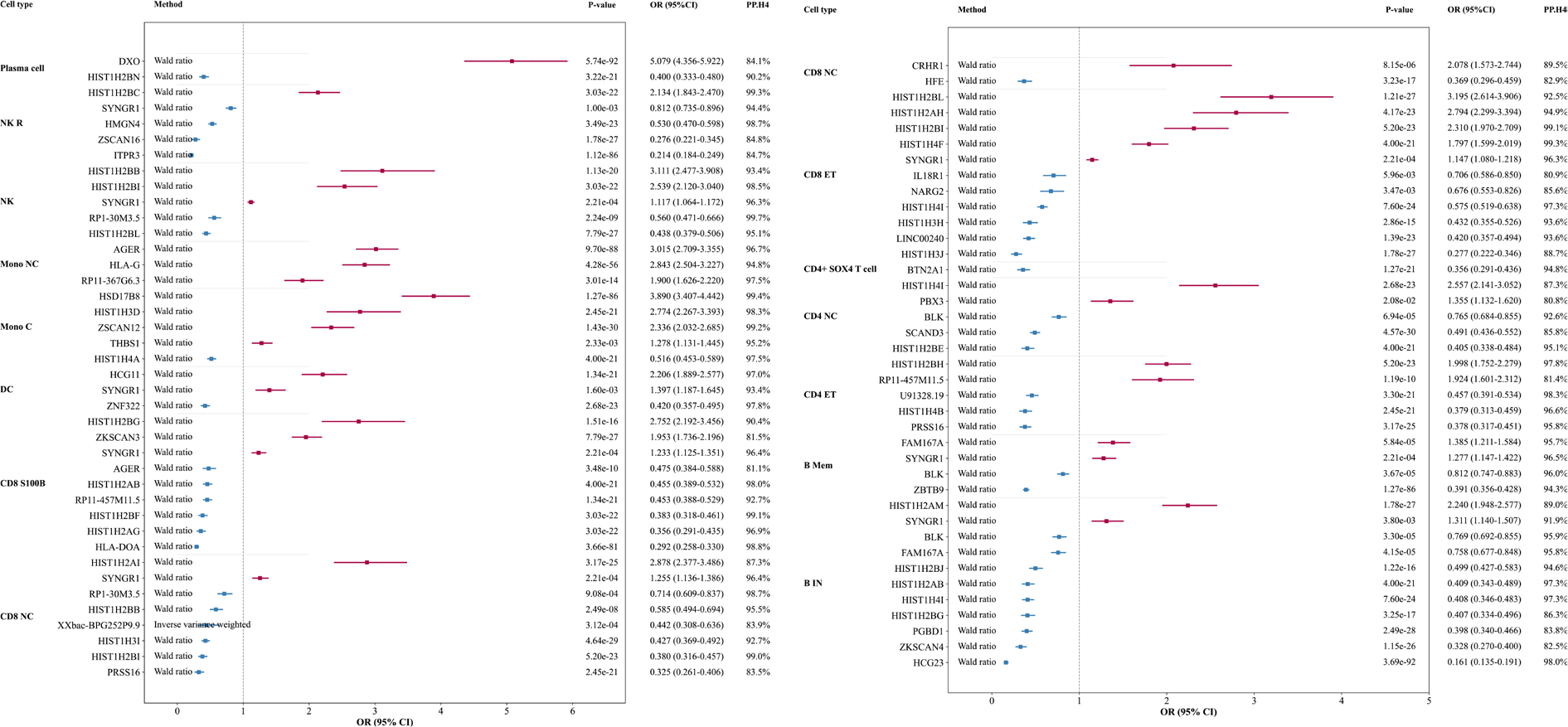
Results: Using a stringent Bayesian colocalization threshold (PP.H4 > 80%) together with significant MR evidence (P < 0.05), we identified multiple immune cell–specific eGenes robustly associated with SS risk across B-cell, T-cell, monocyte, dendritic-cell, natural killer (NK)-cell, and plasma-cell subsets (Figure 1). In immature and naïve B cells, BLK and FAM167A were protective (OR 0.769 and 0.758; PP.H4 around 95.8%), whereas SYNGR1 increased SS risk (OR 1.311; PP.H4 91.9%). Multiple histone-cluster genes showed strong colocalization with both protective and risk-increasing effects, including HIST1H2AM (OR 2.240; PP.H4 89.0%) and several protective HIST1H2/H4 signals (e.g., HIST1H4I, OR 0.408; PP.H4 97.3%). In memory B cells, BLK remained protective (OR 0.812; PP.H4 96.0%), while FAM167A showed a risk-increasing association (OR 1.385; PP.H4 95.7%), highlighting context-dependent regulation within the B-cell lineage. In T-cell subsets, robust associations were observed in both CD4 and CD8 compartments, including risk-increasing signals (e.g., PBX3 in CD4+ naïve and central memory T cells, OR 1.355; PP.H4 80.8%) and multiple strongly colocalized histone-cluster associations (e.g., HIST1H4I in CD4+ naïve and central memory T cells, OR 2.557; PP.H4 87.3%). SYNGR1 showed consistent risk-increasing effects across several T-cell contexts (CD8+ effector memory T cells, OR 1.147; CD8+ naïve and central memory T cells, OR 1.255; PP.H4 ≥ 96.3%). Beyond lymphocytes, dendritic cells showed both risk and protective associations, including HCG11 (OR 2.206; PP.H4 97.0%) and ZNF322 (OR 0.420; PP.H4 97.8%). In classical monocytes, strong risk signals included HSD17B8 (OR 3.890; PP.H4 99.4%) and ZSCAN12 (OR 2.336; PP.H4 99.2%), while non-classical monocytes highlighted AGER (OR 3.015; PP.H4 96.7%) and HLA-G (OR 2.843; PP.H4 94.8%). Notably, AGER showed an opposite, protective association in CD8+ S100B T cells (OR 0.475; PP.H4 81.1%), suggesting cell-type–dependent directionality. In NK cells and NK recruiting cells, several strong signals were observed, including protective effects for ITPR3 (OR 0.214; PP.H4 84.7%) and recurrent SYNGR1 risk associations (PP.H4 ≥ 94.4%). Plasma cells showed the largest effect size for DXO (OR 5.079; PP.H4 84.1%).
Conclusions: By integrating single-cell cis-eQTL with two-sample MR and stringent Bayesian colocalization, we prioritized immune-cell–specific regulatory genes with robust evidence of shared causal variants influencing SS risk. The results indicate both cross-lineage and cell-type restricted mechanisms, highlighted by recurrent B-lineage regulation, widespread risk signals for SYNGR1 across multiple immune contexts, and strong innate and plasma-cell associations. This framework refines the cell-type-resolved genetic architecture of SS and provides a scalable approach for causal gene prioritization using single-cell regulatory genomics.
Forest plot of significant eGenes with shared causal signals for gene expression and Sjögren’s syndrome.
PP.H4 represents the posterior probability that the gene-expression and Sjögren’s syndrome association signals are driven by the same shared causal variant.
CD4 NC, CD4+ Naïve and Central Memory T cell; CD4 ET, CD4+ Effector memory and central memory T cell; B Mem, Memory B cell; B IN, Immature and Naïve B cell; CD8 ET, CD8+ Effector memory T cell; CD8 NC, CD8+ Naïve and Central memory T cell; CD8 S100B, CD8+ S100B T cell; DC, Dendritic cell; Mono C, Classical Monocyte; Mono NC, Non-Classical Monocyte; NK, Natural killer cell; NK R, Natural killer cell Recruiting.
REFERENCES: [1] Yazar S, Alquicira-Hernandez J, Wing K, Senabouth A, Gordon MG, Andersen S, Lu Q, Rowson A, Taylor TRP, Clarke L et al: Single-cell eQTL mapping identifies cell type-specific genetic control of autoimmune disease. Science 2022, 376(6589):eabf3041.
Acknowledgments: NIL.
Disclosure of Interests: None declared.