fetching data ... 
Background: Systemic lupus erythematosus (SLE) is characterized by chronic activation of type I interferon (IFN) signaling, which drives dysregulated T- and B-cell responses and contributes to multisystem tissue damage. The pivotal role of B cells in SLE pathogenesis is underscored by the remarkable efficacy of chimeric antigen receptor T-cell (CAR-T) therapy. Previous studies have reported enrichment of genetic risk variants, skewed V(D)J gene usage, and repertoire abnormalities particularly in naïve and IgM + memory B cells [1]. Although longer complementarity determining region 3 (CDR3) length is generally associated with autoreactivity and polyreactivity, paradoxically [2], CDR3 length in SLE has been reported to be shorter than in healthy individuals [1]. Anifrolumab (ANI), a monoclonal antibody targeting type I IFN receptor subunit 1, has been shown to reduce type I IFN stimulated genes (ISG) expression and disease activity at 3 months; however, its cell type specific immunological effects remain incompletely understood.
Objectives: To delineate the cellular and molecular effects of ANI on immune cell subsets in SLE, with a particular focus on B cell receptor (BCR) repertoire dynamics, using single-cell RNA-seq and BCR-seq of peripheral blood.
Methods: Patients fulfilling the 2019 EULAR/ACR classification criteria for SLE and deemed treatment refractory due to persistent disease activity or difficulty tapering glucocorticoids (GC) were enrolled prior to initiation of ANI therapy. Peripheral blood samples were obtained at baseline and at 3 and 6 months after treatment initiation. CD45 + and CD19 + cells were isolated, followed by single cell partitioning using the 10x Chromium platform. Libraries including BCR sequencing were generated and sequenced, with reads mapped to the human reference genome using Cell Ranger (v7.0.1). Downstream analyses were performed in R (v4.3.2) using Seurat (v4.3.0). ISG scores were calculated as the mean Z-score of IFI27 , IFI44 , IFI44L and RSAD2 . Paired comparisons were conducted using the Wilcoxon signed-rank test. BCR-seq were performed using the Immcantation framework, and generalized linear mixed-effects models adjusted for GC dose were applied.
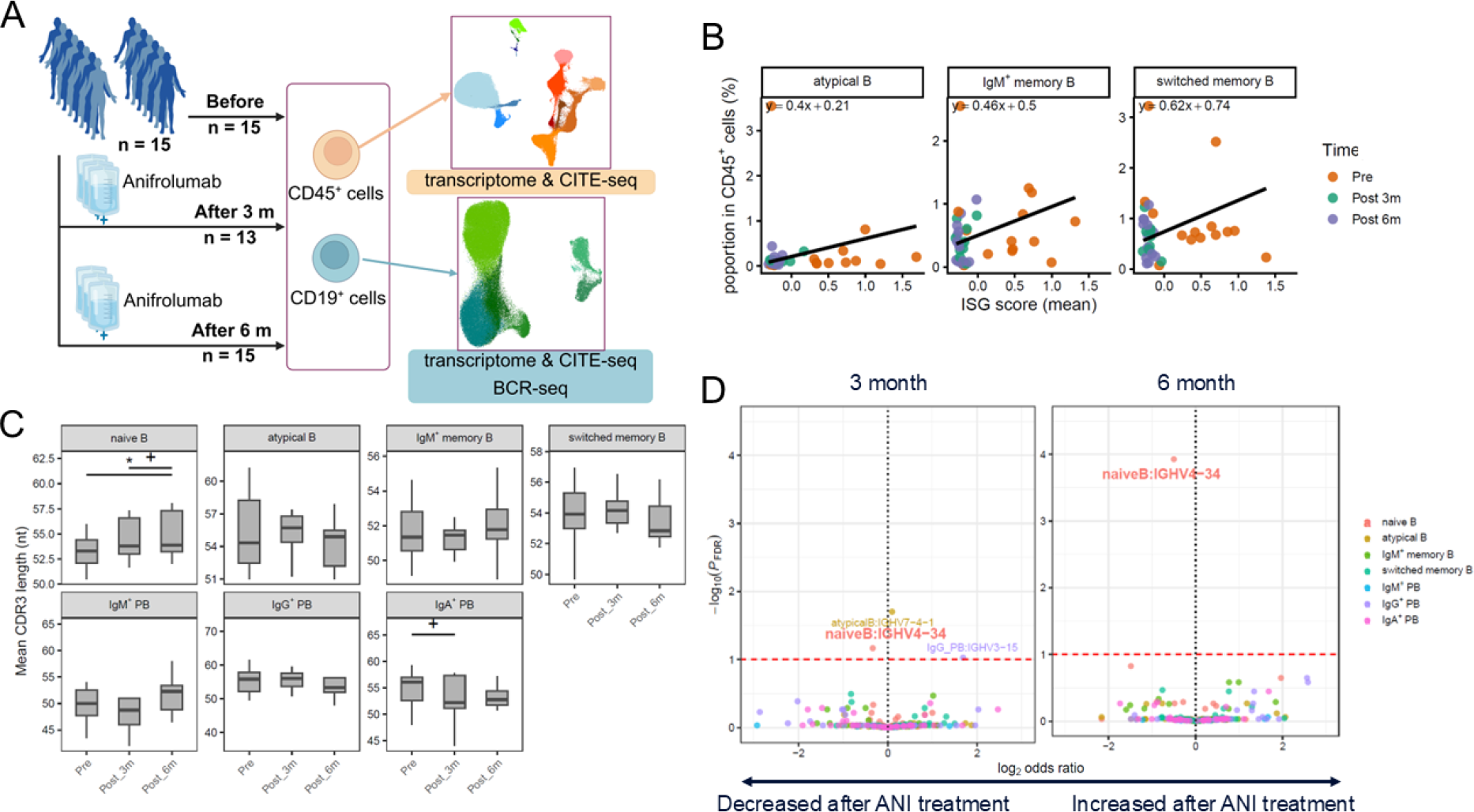
Results: Fifteen patients were enrolled, including fourteen females, with a mean age of 40.3 ± 11.1 years, a mean prednisolone equivalent dose of 6.3 ± 4.7 mg/day, and a mean SLE Disease Activity Index 2000 score of 4.4 ± 2.2 at baseline. ISG scores were reduced across all B cell subsets by 3 months after treatment initiation ( p < 0.05), with no further significant changes observed between 3 and 6 months. Within the CD45 + cells, the proportions of non-classical monocytes, atypical B cells, IgM + memory B cells, and switched memory B cells (SMB) were significantly reduced at 6 months ( p < 0.05) (Figure 1A). BCR sequencing demonstrated elongation of CDR3 length in naïve B cells at 6 months ( p < 0.05, Figure 1B), accompanied by a reduction in IGHV4-34 usage (FDR-adjusted p < 0.1, Figure 1C). IGHV4-34 preferentially paired with IGKV3-20, and the frequency of this pairing was significantly reduced at 6 months ( p < 0.05).
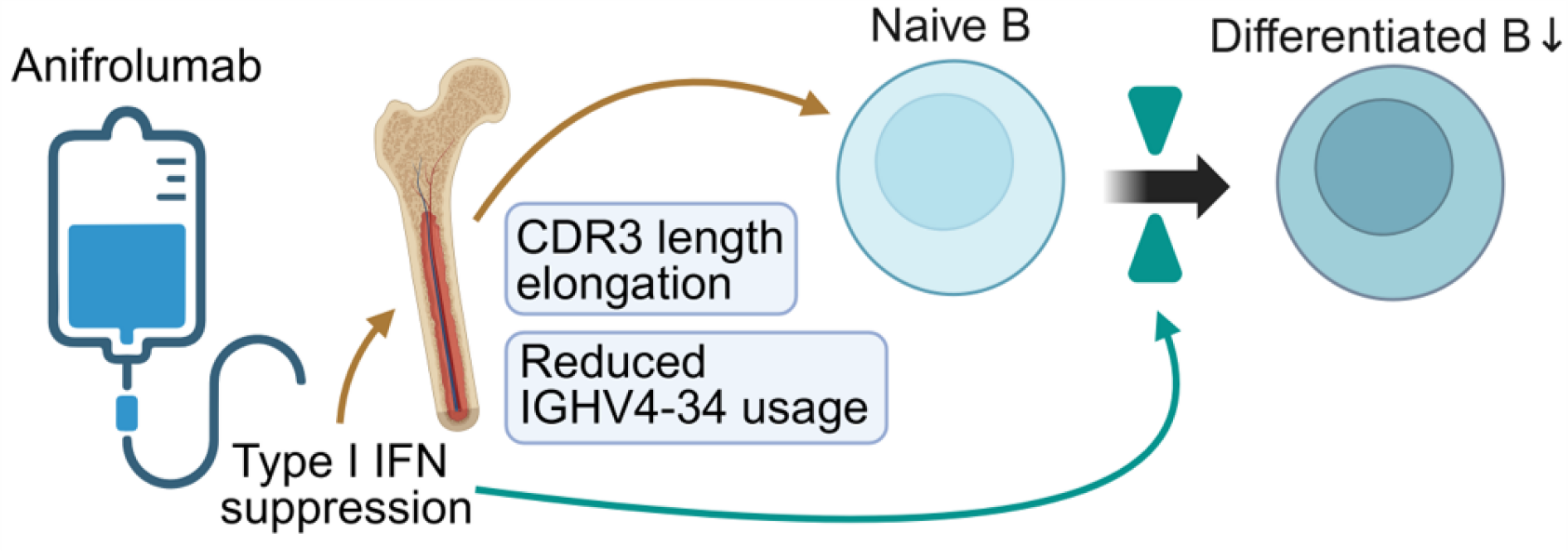
Conclusions: IGHV4-34 is known to exhibit intrinsic autoreactive potential mediated by framework region motifs rather than CDR3-dependent antigen recognition [3]. In naïve B cells, ANI treatment induced an early reduction in ISG scores at 3 months, followed by elongation of CDR3 length and reduced usage of the autoreactivity-associated IGHV4-34 gene at 6 months. These temporal pattern of changes suggests that, within six months after ISG suppression, BCR repertoire remodeling in naïve B cells predominantly reflects correction of central immune tolerance, likely through modulation of V(D)J recombination and junctional diversification during early B cell development. In parallel, decreased proportions of atypical B cells, IgM + memory B cells, and SMB suggest a concomitant reduction in the overall burden of autoreactive B cells (Figure 2).
. Interferon (IFN) suppression associated changes in B cell composition and B cell receptor repertoire following anifrolumab (ANI) treatment. (A) Conceptual overview of the study design. (B) By integrating B cell proportions within CD45 + cells with subset frequencies within CD19 + cells, we estimated B cell subset abundance in the CD45 + compartment and examined its association with type I IFN stimulated genes (ISG) scores. For each B-cell subset, ISG scores and subset proportions from individual patients at each time point were plotted. To illustrate the overall directional change in this relationship over time, a line connecting the mean values at baseline and at 6 months across patients was drawn. (C) Changes in complementarity determining region 3 (CDR3) length, measured in nucleotides (nt), were assessed across B cells based on patient level mean values across time points (Wilcoxon signed rank test * p < 0.05, + p < 0.1). (D) Changes in IGHV gene combinations across B cell subsets were assessed using generalized linear mixed models including glucocorticoid dose as a covariate, comparing baseline with 3 months (left) and 6 months (right) after treatment (FDR-adjusted p < 0.1; labeled, with decreases after ANI highlighted in bold and larger text).
Anifrolumab (ANI) mediated suppression of type I interferon (IFN) signaling was associated with a remodeling of the B-cell receptor (BCR) repertoire. ANI mediated suppression of type I IFN signaling was associated with elongation of complementarity determining region 3 (CDR3) length and reduced usage of the IGHV4-34 gene in naïve B cells, likely reflecting altered V(D)J recombination and junctional diversification in the bone marrow. In parallel, type I IFN suppression was associated with decreased proportions of differentiated B cell subsets, including atypical B cells, IgM + memory B cells, and switched memory B cells.
REFERENCES: [1] Ota M. Ann Rheum Dis 2023;82:1455-1463.
[2] Aguilera I. Immunology 2001;102:273-80.
[3] Deguine J. J Exp Med 2024;221:e20231314.
Acknowledgments: NIL.
Disclosure of Interests: Keigo Terada: None declared, Yumi Tsuchida AbbVie, Asahi Kasei Corporation, Astellas Pharma, AstraZeneca, AYUMI Pharmaceutical Co, Daiichi Sankyo Company, Eisai Co, Eli Lilly, GlaxoSmithKline, Janssen Pharmaceutical, Nippon Shinyaku Co, Novartis Pharmaceuticals, Otsuka Pharmaceutical, Taisho Pharmaceutical Co, and UCB., Toshiyuki S Ushijima: None declared, Yuichi Suwa Abbvie., Kazusa Saegusa: None declared, Saeko Yamada: None declared, Mineto Ota MSD, AstraZeneca, and Chugai Pharmaceutical., GlaxoSmithKline, the Mochida Memorial Foundation for Medical and Pharmaceutical Research, the Astellas Foundation for Research on Metabolic Disorders, the Chugai Foundation for Innovative Drug Discovery Science, and affiliation with the University of Tokyo Social Cooperation Program funded by Chugai Pharmaceutical Co., Ltd., Tomohisa Okamura Asahi Kasei Corporation, and Bristol-Myers Squibb Company, Affiliation with the University of Tokyo Social Cooperation Program funded by Chugai Pharmaceutical Co., Ltd., Keishi Fujio Asahi Kasei Pharma., Chugai, Abbvie, Asahi Kasei Pharma, Bristol Myers Squibb, AstraZeneca, Mitsubishi Tanabe, Eisai, Gilead, Eli Lilly, Pfizer, Taisho, Astellas, Daiichi Sankyo, Novartis, GlaxoSmithKline, and Alexion Pharma., Asahi Kasei Pharma., Chugai, Abbvie, Asahi Kasei Pharma, Bristol Myers Squibb, AstraZeneca, Eisai, Tsumura, and Taisho.