fetching data ... 
Background: Systemic sclerosis (SSc) is a severe autoimmune disease with multiorgan involvement. Current therapeutic strategies remain organ-specific, lacking systemic approaches. Recent single-cell analysis of SSc synovium revealed a robust type I and type II interferon (IFN) signature across major cellular populations, including FOSB+ fibroblasts, venous endothelial cells, and infiltrating macrophages, as well as activation of the alternative complement pathway in fibroblasts [1]. However, it remains unknown whether these signatures are shared across multiple affected organs and could thus represent systemic targets.
Objectives: We aimed to determine whether (i) IFN and alternative complement pathway activation represent a systemic molecular hallmark of SSc and (ii) whether these signatures originate from shared pathogenic cellular subpopulations in lung and skin.
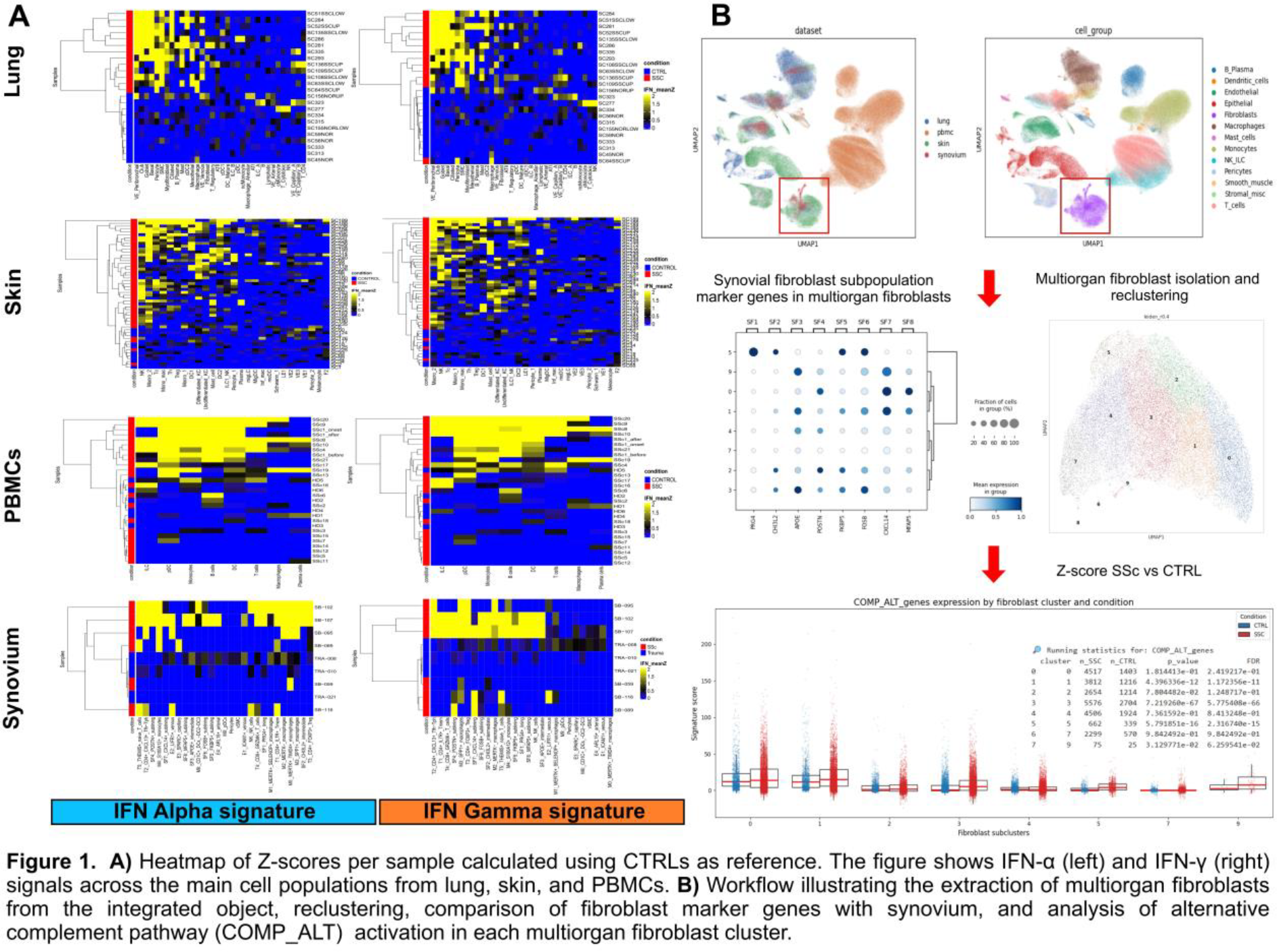
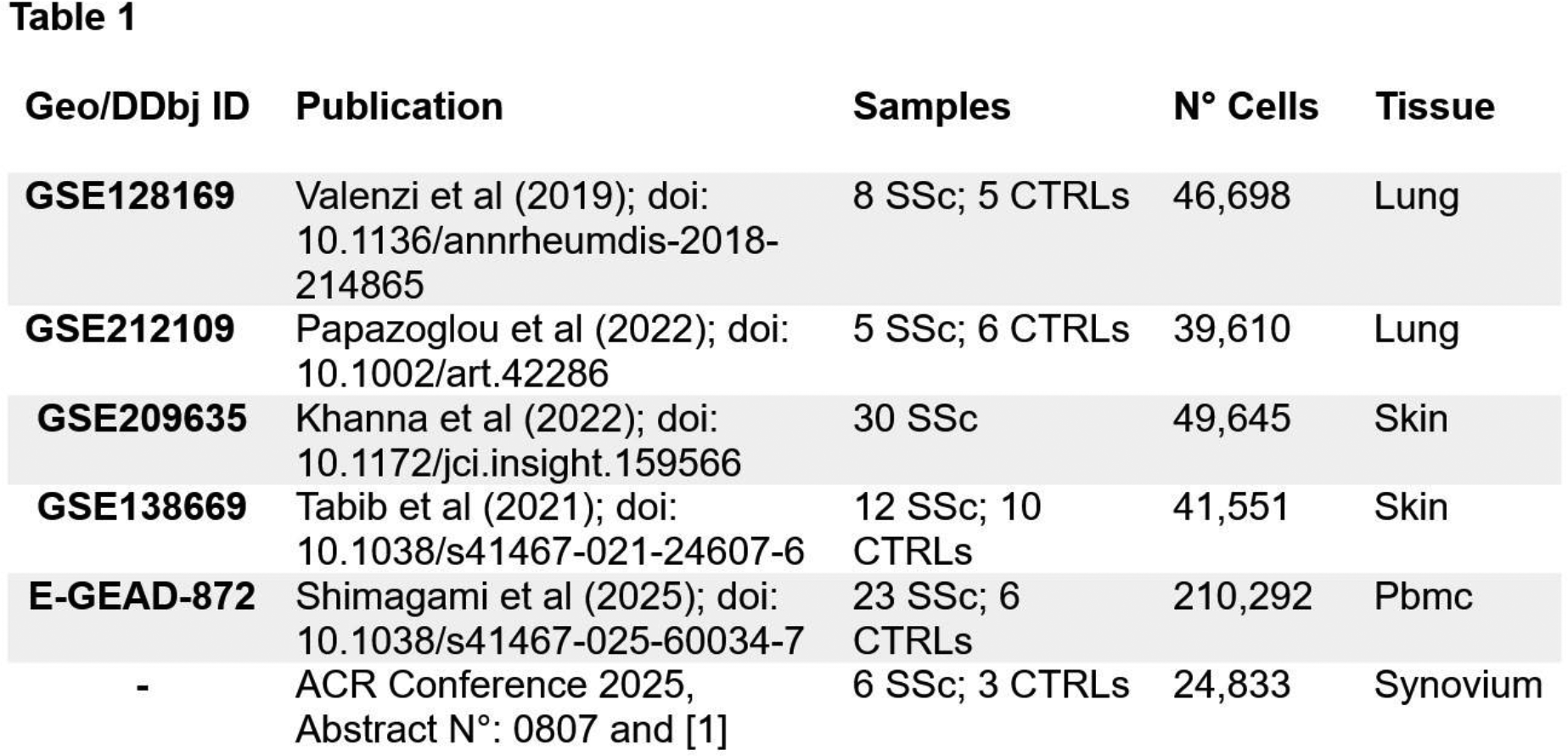
Methods: We compiled 5 publicly available SSc single-cell datasets from skin, lungs, PBMC, and 1 in-house dataset from synovium ( Table 1 ) [1]; each was then processed and normalized independently by Scanpy (v1.11). The cells were annotated using Celltypist models, followed by differential gene expression (pydeseq2) and gene set enrichment analysis (GSEA) comparing SSc with controls (CTRLs). Fibroblast, monocyte-macrophage, and endothelial populations from all tissues were reintegrated and reclustered (Leiden resolution = 0.4) to identify pathogenic states shared with synovium. Type I and II IFN and the alternative complement pathway activation from Hallmark gene sets were quantified by gene-level Z-scores relative to controls and validated by hierarchical clustering (Euclidean, complete linkage).
Results: After quality control filtering, a total of 86,563 lung cells from 24 patients (13 interstitial lung disease SSc; 11 controls), 91,541 skin cells from 52 patients (46 diffuse cutaneus SSc; 6 controls), 210,881 PBMCs from 29 patients (23 SSc; 6 controls), and 24,833 synovium cells from 9 patients (6 SSc; 3 CTRLs) were included. Multi-organ integration with the synovium dataset revealed overlapping fibroblast, endothelial, and immune populations across skin, lung, and synovium, while PBMCs aligned only within immune compartments. In individual tissues and in the integrated atlas, type I and II IFN and alternative complement signatures consistently distinguished SSc from controls. This was across all major cell types, as shown in Figure 1A for IFN in individual tissues and in Figure 1B for alternative complement in multiorgan fibroblasts. As in synovium, venous endothelial cells and fibroblasts in the lung, as well as macrophages in the skin, showed the most prominent IFN-driven transcriptional activation. Fibroblast: Reclustering of individual tissues identified seven lung fibroblast subclusters (L1–L7) and five skin subclusters (S1–S5) from 4,273 lung and 22,780 skin fibroblasts, respectively. In the lung, a distinct fibroblast population (L3) showed strong co-activation of type I and II IFN signaling together with alternative complement pathways, resembling the pathogenic fibroblast phenotype previously described in SSc synovium. This population also displayed elevated TGF-β and Wnt signaling and a hypermetabolic profile characterized by increased oxidative phosphorylation, amino-acid, and glucose utilization. In skin, disease–control separation was less pronounced, although clusters S1 and S2 were enriched for inflammatory-response pathways, with IFN-II activation largely confined to S1. Cross-organ integration of fibroblasts from lung, skin, and synovium identified ten multi-organ clusters (M0–M9). M0 and M7 clusters were composed of skin fibroblasts and enriched for S1 and S2 cells, respectively, suggesting distinct inflammatory programs within skin. In contrast, the IFN-driven lung cluster L3 mapped to the shared multi-organ cluster M3, which also included IFN-activated FOSB + and APOE + synovial fibroblasts and showed significantly higher IFN-I and IFN-II signature scores in SSc compared with controls. The other clusters displayed a homogeneous contribution from lung, skin, and synovium fibroblasts, supporting shared pathogenic programs, whereas M5 was synovium-specific lining fibroblasts. M6 and M8 were excluded as small clusters of non-fibroblast cells. Macrophage : In skin, two macrophage clusters (S1 and S2) discriminated SSc from controls and displayed strong IFN-I, IFN-II, and NF-κB activation, similar to synovial macrophages. Integration confirmed that skin S2 macrophages colocalized with synovial M2 (MERTK − ), M3 (SPP1 + ), and M4 (S100A12 + ) monocyte subsets, forming two IFN-high multi-organ clusters. PBMC analysis stratified SSc patients into IFN I and II positive (SSc IFN + ) and IFN I and II negative (SSc IFN − ) groups, primarily driven by monocytes and plasmacytoid dendritic cells. Notably, only monocytes recapitulated complement-related activation, indicating partial overlap with tissue inflammatory programs. Endothelial : Given the strong IFN activation previously observed in synovial endothelial cells, we next assessed endothelial states in lung and skin. Lung, but not skin endothelial cells contained a distinct IFN-I/II–high subset (L2) that clearly separated SSc from controls and colocalized with the ICAM1 + (E1) and ARL15 + (E4) synovial endothelial subsets sharing a common IFN-driven phenotype.
Conclusions: This multi-organ analysis revealed a shared type I and II IFN and alternative complement pathway activation signature across multiple organs in SSc and identified shared multi-organ fibroblast, endothelial, and macrophage-monocytes populations. Importantly, we identify a shared fibroblast state with high metabolic activity and strong co-activation of IFN and alternative complement pathways. These findings support a systemic pathogenic program in SSc and position IFN and alternative complement-targeted approaches as promising candidates for systemic therapeutic intervention in SSc.
REFERENCES: [1] C. Geiss, et al OP0192. Ann Rheum Dis, 83;48 (2024)
Acknowledgments: NIL.
Disclosure of Interests: Cristian Iperi: None declared, Celina Geiss: None declared, Philip Stauffer: None declared, Anna-Maria Hoffmann-Vold Boehringer Ingelheim, Janssen, Medscape, Merck Sharp & Dohme, Novartis, Roche, AbbVie, Avalyn, Astra Zeneca, Boehringer Ingelheim, Bristol Myers Squibb, Calluna Pharma, Genentech, Janssen, Medscape, Merck Sharp & Dohme, Pliant, Roche, Werfen, Astra Zeneca, Boehringer Ingelheim, Janssen, Oliver Distler Boehringer Ingelheim, 4P-Pharma, Abbvie, Acepodia, Aera, AnaMar, Anaveon, Argenx, AstraZeneca, Avalyn, Boehringer Ingelheim, BMS, Calluna, Cantargia, CSL Behring, EMD Serono, Galderma, Galapagos, Gossamer, Hemetron, Innovaderm, Kali, Lilly, Mediar, MSD Merck, Nkarta, Novartis, Oorja Bio, Orion, Pliant, Prometheus, Quell, Scleroderma Research Foundation, Skyhawk, Tandem, Topadur, UCB and Umlaut.bio., Kymera, Mitsubishi Tanabe, UCB, Caroline Ospelt: None declared, Muriel Elhai Boehringer Ingelheim, Vontobel Stiftung, Pfizer, Novartis Foundation for Bio-Medical Research, Iten Kohaut foundation, Kurt und Senta Herrmann foundation, Foundation for Research in Rheumatology (FOREUM), University Zurich, Janssen, Walter and Gertrud Siegenthaler Foundation.