fetching data ... 
Background: Selective inhibition of PDE4B has very recently been shown to be effective for progressive fibrosing interstitial lung diseases (ILDs) in two large randomized controlled clinical trials. With its fibro-immunomodulatory effects, PDE4B inhibition could be a promising treatment strategy for systemic sclerosis (SSc). However, cell type–specific expression patterns across affected tissues in SSc have not been systematically examined.
Objectives: To investigate whether PDE4B expression is consistently dysregulated in blood and across the fibrotic skin and lung, supporting its potential as a disease-relevant target for pharmacological intervention for SSc.
Methods: For the lung, two publicly available single-cell RNA-sequencing (scRNA-seq) datasets were integrated: one with SSc–associated interstitial lung disease (SSc-ILD, n = 4, GSE128169) samples and one with idiopathic pulmonary fibrosis (IPF, n = 4, GSE128033) samples. Heathy control (HC) lung (n=4) samples were also included (n = 4, GSE128169). In addition, we performed scRNA-seq on peripheral blood mononuclear cells (PBMCs) from 25 early and active SSc patients and 9 age-matched HCs using the Chromium Next GEM protocol (10X Genomics). All transcriptomic analyses were performed using Seurat (v5.1.0). Cells were clustered and subsequently manually annotated for the lungs and annotated using Azimuth for the PBMCs. Differential expression analysis was performed using MAST . To define significant cell types, a |log2FC| > 0.5 was used as a cut-off. Additional analyses were performed in skin scRNA-seq data using an SSc cell atlas that integrated publicly available skin scRNA-seq datasets. From this, two skin scRNA-seq datasets were downloaded and normalized (GSE138669, 12 SSc and 10 HC; GSE249279, 22 SSc, 18 HC). The differential expression analysis results were used. Normalized PDE4B expression from bulk RNA-seq of skin from early, diffuse cutaneous SSc patients (N = 48) enrolled in the Prospective Registry for Early Systemic Sclerosis (PRESS) were compared to HCs (N = 33). Skin biopsies (9 SSc, 4 healthy) were formalin-fixed and paraffin-embedded for immunohistochemistry (PDE4B) and subsequently analysed using QuPath to measure the percentage of positive cells.
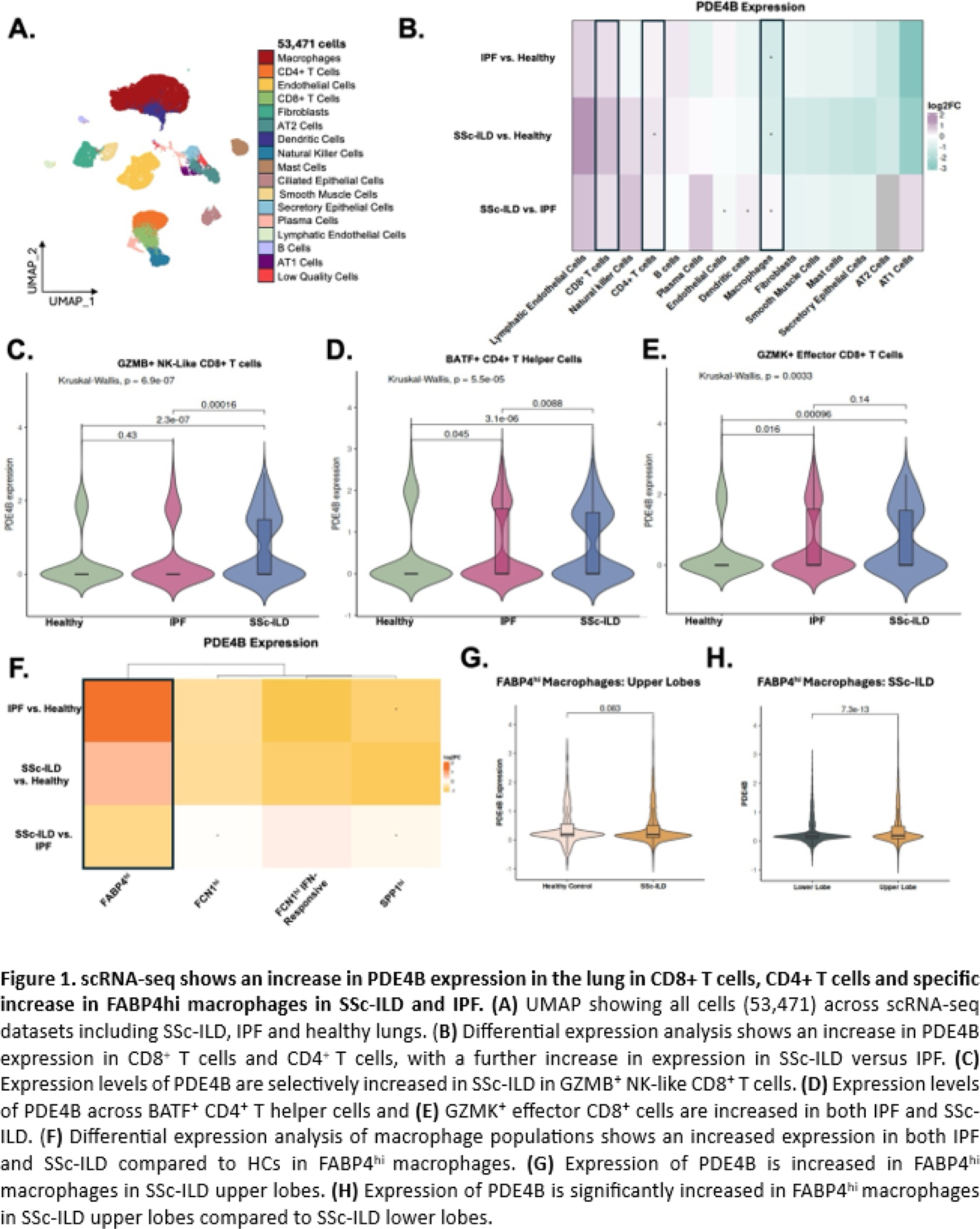
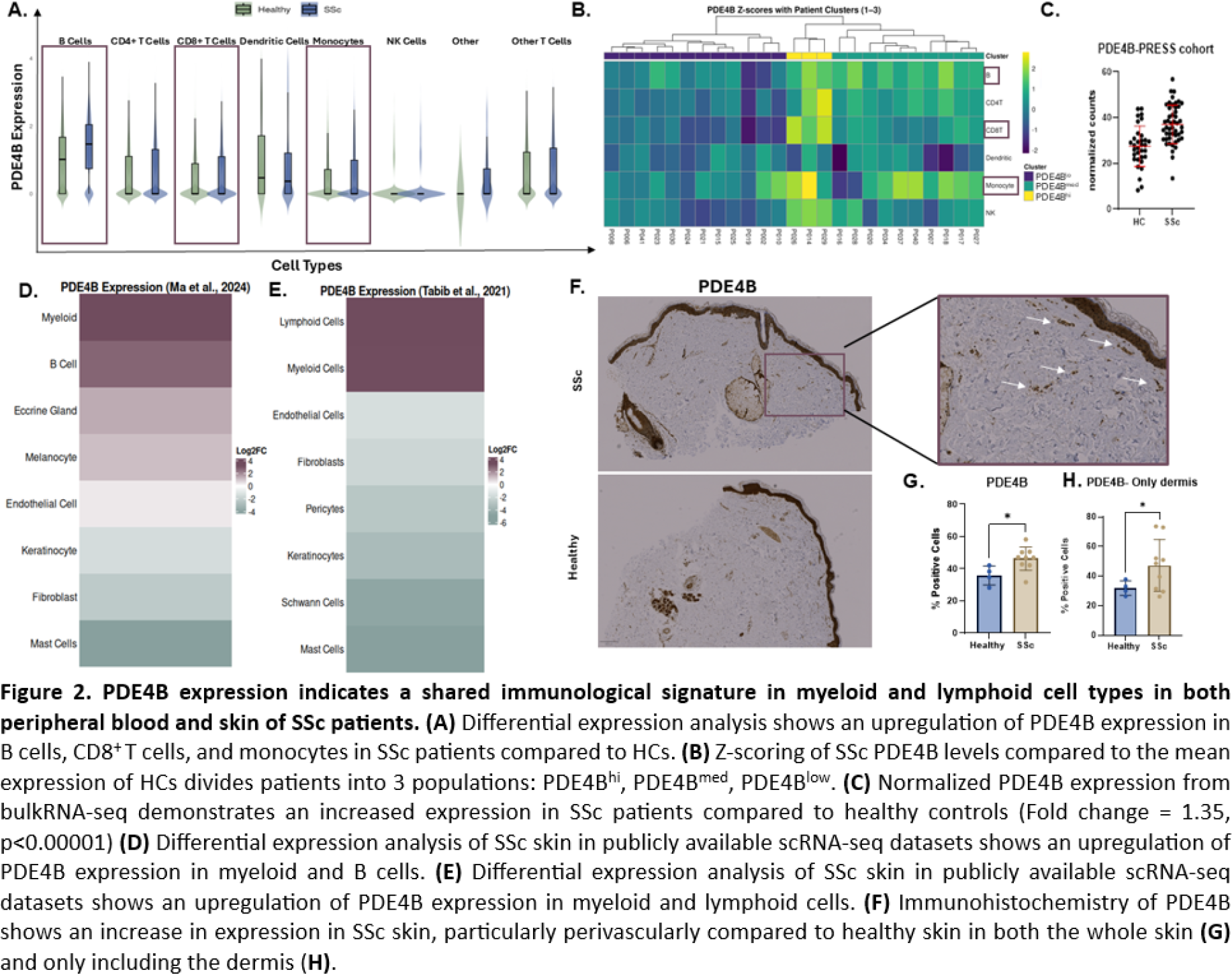
Results: For the lung, 53,471 cells were retained after quality control (Figure 1A ). Differential expression analysis showed an increase in PDE4B expression in several cell types, including both CD8 + and CD4 + T cells in IPF and SSc-ILD compared with HCs (Figure 1B ). Furthermore, there was an additional upregulation in SSc-ILD relative to IPF (Figure 1B ). To assess whether T cell populations exhibit homogeneous PDE4B expression, the T cell compartment was further divided into five subpopulations: BATF + CD4 + T helper, CCR7 + CD4 + naïve/central memory, GZMB + NK-like CD8 + , GZMK + effector CD8 + , and activated/exhausted T reg -like CD4 + T cells. GZMK + CD8 + effector T cells had increased PDE4B expression in both IPF and SSc-ILD relative to healthy lungs, whereas GZMB + CD8 + T cells displayed an SSc-ILD–specific increase. BATF + CD4 + T helper cells also demonstrated elevated PDE4B in both SSc-ILD and IPF compared with HCs (Figure 1C-E ). Interestingly, fibroblasts showed an overall negative fold-change in the disease-conditions compared to HCs, thus placing an additional emphasis on the immune component in fibrosing ILDs (Figure 1B ). Lung macrophages globally demonstrated decreased PDE4B expression in both IPF and SSc-ILD relative to healthy tissue (Figure 1B ). However, when we further divided macrophages into the five known subpopulations: SPP1 hi , FCN1 hi inflammatory, FCN1 hi IFN-responsive, FABP4 hi alveolar, and a SPP1 hi /FABP4 hi mixed population, FABP4 hi macrophages had an overall increase in PDE4B expression in IPF and SSc-ILD compared with healthy lungs (Figure 1F ). When comparing the upper and lower lobes, FABP4 hi macrophages in SSc-ILD showed a significant increase in PDE4B expression in the upper lobes compared to HCs (Figure 1G ). Additionally, in SSc-ILD the upper lobe demonstrated increased PDE4B expression compared to the lower lobe (Figure 1H ). In PBMCs, PDE4B expression was significantly increased in B cells, CD8 + T cells, and monocytes of SSc patients (Figure 2A ). Hierarchical clustering divided patients into 3 groups showing PDE4B lo , PDE4B med and PDE4B hi expression patterns across cell types (Figure 2B ). Next, we aimed to identify clinical parameters associated with high PDE4B expression in PBMCs allowing enrichment for clinical trials. However, using different approaches including linear regression analyses of the Z-score per each cell type with continuous clinical variables (including mRSS, FVC, rEUSTAR-AI, and SCTC-AI) we could not identify relevant associations. PDE4B expression is globally increased in SSc compared to age- and gender-matched HCs ( Figure 2C ). To identify the cellular sources underlying this global increase, differential expression analysis of scRNA-seq data from SSc skin was used. This revealed upregulation of PDE4B in both myeloid and lymphoid populations (Figure 2D-E ). Immunohistochemistry confirmed increased PDE4B protein levels in SSc skin compared with HCs (Figure 2F ). Interestingly, PDE4B expression was prominent in the epidermis, around glandular structures and blood vessels, and within perivascular regions. To determine whether the observed increase was driven by epidermal expansion in SSc skin, we restricted the analysis to the dermis. Despite this, PDE4B expression remained significantly elevated in SSc samples, indicating that increased expression is not solely attributable to epidermal thickening (Figure 2G-H ).
Conclusions: PDE4B is selectively upregulated across key immune cell populations across SSc organs and in the peripheral blood. Immune cells appear to be the primary target for PDE4B inhibition in an immune-fibrotic disease like SSc. Overall, PDE4B expression emerges as a shared immunologic feature across SSc tissues, supporting its relevance as a potential therapeutic target.
REFERENCES: NIL.
Acknowledgments: NIL.
Disclosure of Interests: Astrid Hofman: None declared, Pietro Bearzi: None declared, Laura Much: None declared, Maryam Asadikorayem: None declared, Andrea Laimbacher: None declared, Helen Jarnagin: None declared, Zhiyung Gong: None declared, Michael Whitfield Abbvie, Boehringer Ingelheim, Bristol-Myers Squibb, Celdara Medical, LLC, UCB Biopharma, Abbvie, Boehringer Ingelheim, Bristol-Myers Squibb, Celdara Medical, LLC, UCB Biopharma, Abbvie, Boehringer Ingelheim, Bristol-Myers Squibb, Celdara Medical, LLC, UCB Biopharma, Shervin Assassi Boehringer Ingelheim, Abbvie, aTyr, AstraZeneca, Boehringer Ingelheim, Bymmunity, Candid, CSL Behring, Merck, Mitsubishi Tanabe, Takeda and UCB, Abbvie, aTyr, Boehringer Ingelheim and Janssen, Anna-Maria Hoffmann-Vold Boehringer Ingelheim, Janssen, Medscape, Merck Sharp & Dohme, Novartis, Roche, AbbVie, Avalyn, Astra Zeneca, Boehringer Ingelheim, Bristol Myers Squibb, Calluna Pharma, Genentech, Janssen, Medscape, Merck Sharp & Dohme, Pliant, Roche, Werfen, Astra Zeneca, Boehringer Ingelheim, Janssen, Mike O. Becker Amgen, Vifor, Grant/research support from: Foundation for research in Rheumatology (FOREUM), EMDO Foundation, Novartis foundation for medical-biological research, Innovative Medicines Initiative (IMI)., GSK, Amgen, Novartis, Vifor. Congress support from Vifor., Elena Pachera: None declared, Oliver Distler Boehringer Ingelheim, 4P-Pharma, Abbvie, Acepodia, Aera, AnaMar, Anaveon, Argenx, AstraZeneca, Avalyn, Boehringer Ingelheim, BMS, Calluna, Cantargia, CSL Behring, EMD Serono, Galderma, Galapagos, Gossamer, Hemetron, Innovaderm, Kali, Lilly, Mediar, MSD Merck, Nkarta, Novartis, Oorja Bio, Orion, Pliant, Prometheus, Quell, Scleroderma Research Foundation, Skyhawk, Tandem, Topadur, UCB and Umlaut.bio, Kymera, Mitsubishi Tanabe, UCB.