fetching data ... 
Background: Gouty arthritis (GA) is a recurrent inflammatory disease driven by immune dysregulation following monosodium urate (MSU) crystal deposition. Current therapies often fail to restore long-term immune homeostasis, highlighting the need for novel strategies that can both suppress inflammation and re-establish immune tolerance.
Objectives: To design and evaluate a bioresponsive therapeutic platform for targeted treatment of GA, which combines the immunomodulatory properties of regulatory T cell (Treg)-derived exosomes and growth arrest-specific protein 6 (Gas6), with active targeting to inflamed joints.
Methods: We engineered a hybrid system by co-loading Gas6 onto Treg-derived exosomes. To achieve targeted delivery, the exosomes were modified to leverage CCR2-guided chemotaxis for homing to inflamed joints. The therapeutic efficacy was evaluated in an MSU-induced murine model of GA. Key outcome measures included clinical symptom assessment, analysis of immune cell populations (particularly Tregs and TH17 cells), and evaluation of overall immune homeostasis.
Results: The engineered Treg-exosome/Gas6 platform effectively alleviated key clinical symptoms in the murine GA model. Mechanistically, the treatment promoted the expansion of regulatory T cells while simultaneously suppressing pathogenic TH17 responses. This coordinated immunomodulation led to the restoration of durable immune homeostasis by bridging innate and adaptive immunity and reprogramming the joint tissue environment toward a tolerogenic state.
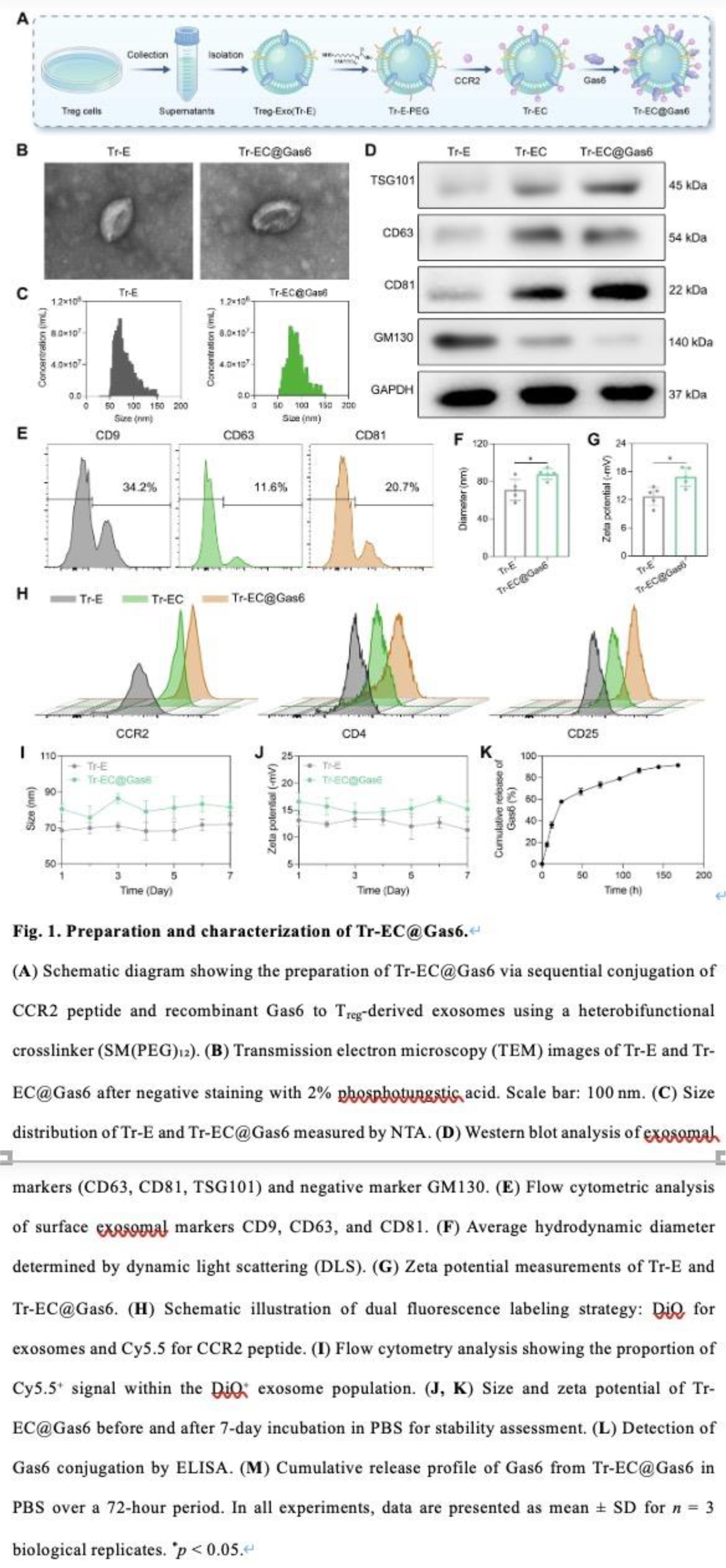
Conclusions: Our study presents a novel, cell-free, and CCR2-guided immunoengineering approach that achieves targeted delivery and synergistic immune reprogramming. This platform represents a promising and effective strategy for treating gouty arthritis and potentially other disorders characterized by immune imbalance. As illustrated in Figure 1A , we constructed dual-functional exosomes (Tr-EC@Gas6) by sequentially conjugating purified T reg -derived exosomes with a CCR2-targeting peptide and recombinant Gas6 protein via a heterobifunctional crosslinker. The modification was carried out under mild stirring at 4 °C and subsequently purified through sucrose density gradient centrifugation to remove unbound components. TEM imaging ( Figure 1B ) revealed that both native exosomes (Tr-E) and engineered exosomes (Tr-EC@Gas6) retained the characteristic cup-shaped morphology typical of exosomes. This indicated that the surface modification process did not compromise the ultrastructural integrity of the vesicles. NTA ( Figure 1C ) showed a rightward shift in particle size distribution following surface functionalization: Tr-E exhibited a hydrodynamic diameter of ~75 nm, while Tr-EC@Gas6 increased to ~90 nm. Western blot analysis ( Figure 1D ) confirmed the enrichment of classical exosomal markers (CD63, CD81, TSG101) in both groups, and the absence of the Golgi marker GM130 excluded cellular contamination. These data verified the exosomal nature of the vesicles and supported the purity of the preparations. Furthermore, quantitative flow cytometric analysis ( Figure 1E ) showed comparable expression levels of CD9, CD63, and CD81, further validating the successful isolation and integrity of the exosomal populations post-modification. Size quantification ( Figure 1F ) showed that the average diameter of Tr-EC@Gas6 (89.4 ± 1.5 nm) was significantly larger than that of Tr-E (74.6 ± 2.7 nm), consistent with NTA results and providing additional support for successful conjugation. The correlation between DLS and NTA reinforces the reliability of the physical characterization data. Zeta potential measurements ( Figure 1G ) revealed no significant change in surface charge between Tr-E (−15.8 ± 2.8 mV) and Tr-EC@Gas6 (−12.4 ± 2.3 mV). As shown in Figure 1I , the percentage of Cy5.5 + events increased significantly from 7.19 ± 2.54% in the Tr-EC group to 82.63 ± 3.74% in the Tr-EC@Gas6 group, representing an approximate 11.5-fold enrichment. As shown in Figure 1J–K , both hydrodynamic diameter and zeta potential remained statistically unchanged, indicating that the dual conjugation process did not affect the structural or physicochemical stability of the exosomes. As shown in Figure 1L , Tr-EC@Gas6 displayed significantly higher Gas6 surface levels compared to Tr-EC and Tr-E, confirming the effective incorporation of Gas6 during the dual-modification process. Furthermore, cumulative release analysis in PBS ( Figure 1M ) demonstrated a sustained release profile over 72 hours, indicating stable surface binding with gradual dissociation under physiological conditions.
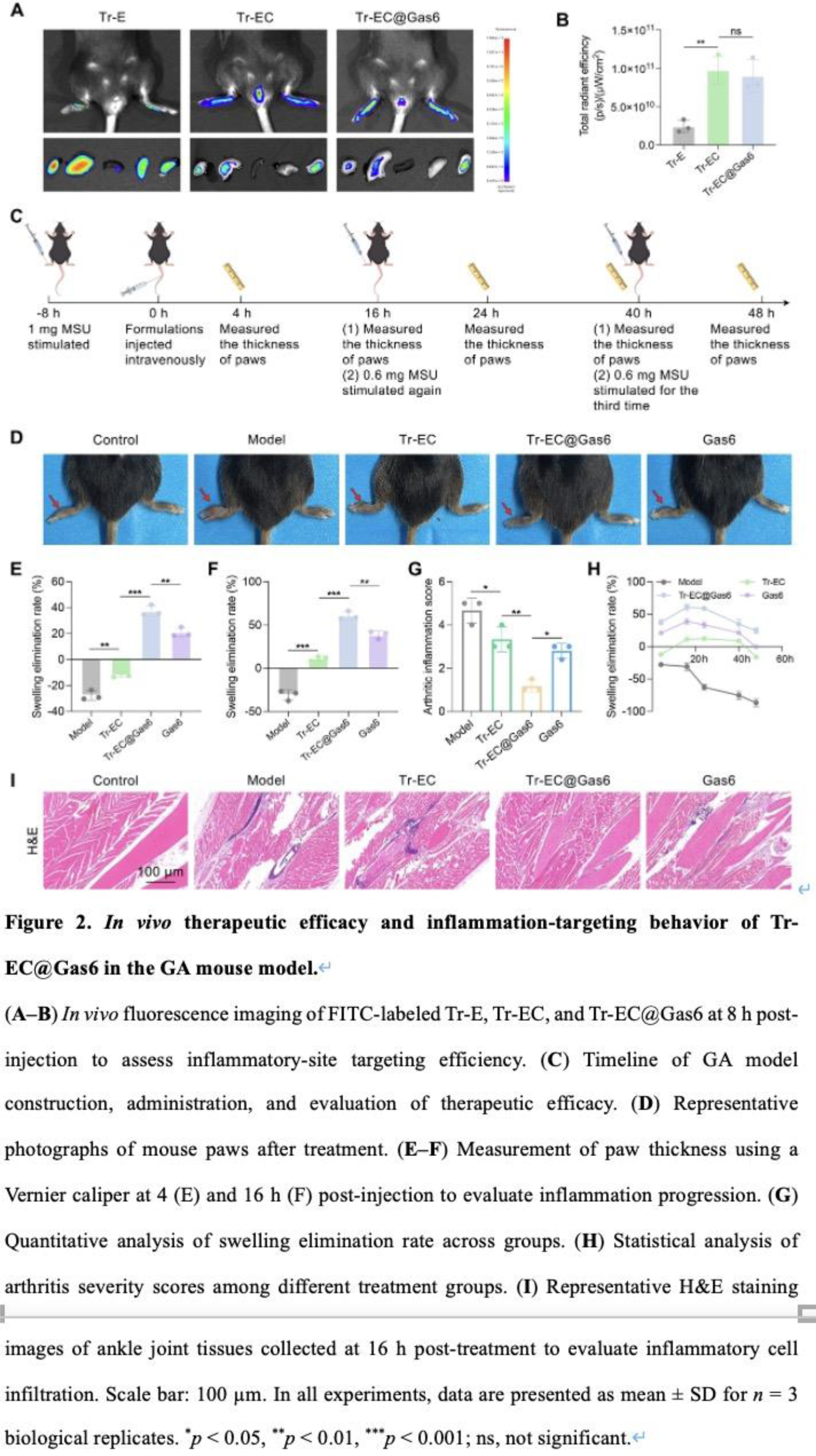
As shown in Figure 2A–B , Tr-EC and Tr-EC@Gas6 exhibited significantly higher fluorescence signals at the ankle joints compared to Tr-E, indicating enhanced targeting to the inflamed sites. Building on the potent anti-inflammatory activity in vitro and the enhanced targeting efficiency in vivo , we further assessed the therapeutic efficacy of Tr-EC@Gas6 in the GA mouse model. As shown in Figure 2C , GA was induced by intra-articular MSU injection 8 hours prior to treatment, resulting in visible swelling of the right hind paw, which peaked at approximately 12 hours. As shown in Figure 2D-E , both the Model and Tr-EC groups exhibited continued swelling at 4 hours, with only a marginal reduction in the latter. At 16 hours post-treatment ( Figure 2F ), Gas6 exhibited moderate efficacy (38.29% reduction), whereas Tr-EC@Gas6 demonstrated a markedly enhanced anti-inflammatory effect, achieving a 69.78% resolution rate. Furthermore, the severity of joint inflammation was evaluated using an arthritis scoring system based on macroscopic imaging ( Figure 2G ). The results were consistent with the swelling analysis, showing that Tr-EC@Gas6 markedly alleviated visible inflammation and joint deformation compared with other groups. To further assess the therapeutic outcomes at the histological level, ankle tissues were harvested 16 hours post-treatment and subjected to H&E staining ( Figure 2I ).
REFERENCES: NIL.
Acknowledgments: NIL.
Disclosure of Interests: None declared.