fetching data ... 
Background: Regression of skin fibrosis is a feature of the natural disease trajectory in systemic sclerosis (SSc). The molecular mechanisms underlying this regression remain unclear. Elucidating these mechanisms might help to identify therapeutic targets for established skin fibrosis.
Objectives: To establish a molecular fingerprint characterizing regression of skin fibrosis in SSc patients.
Methods: All patients met the 2013 ACR/EULAR SSc classification criteria. Skin fibrosis regression/progression was defined by a ≥5-point decrease/increase, respectively, in the modified Rodnan Skin Score (mRSS) at 12-month follow-up. Serum proteomics (Olink High Throughput Explore, 5,400 proteins) was performed on 44 SSc patients, and differentially abundant proteins were identified using limma (FDR < 0.3). Peripheral blood mononuclear cells (PBMCs) single cell RNA-sequencing (scRNA-seq) was generated from 10 SSc patients. In situ spatial transcriptomics was performed on skin using the Xenium 5K platform. Quality control retained boundary-segmented cells containing >15 transcripts and >10 unique genes, with nucleus areas of 6–80 µm 2 . Spatial clustering was performed with BANKSY and integrated using Harmony; cell types were manually annotated and reintegrated using STACAS . All transcriptomic analyses were conducted with Seurat (v5.1.0).
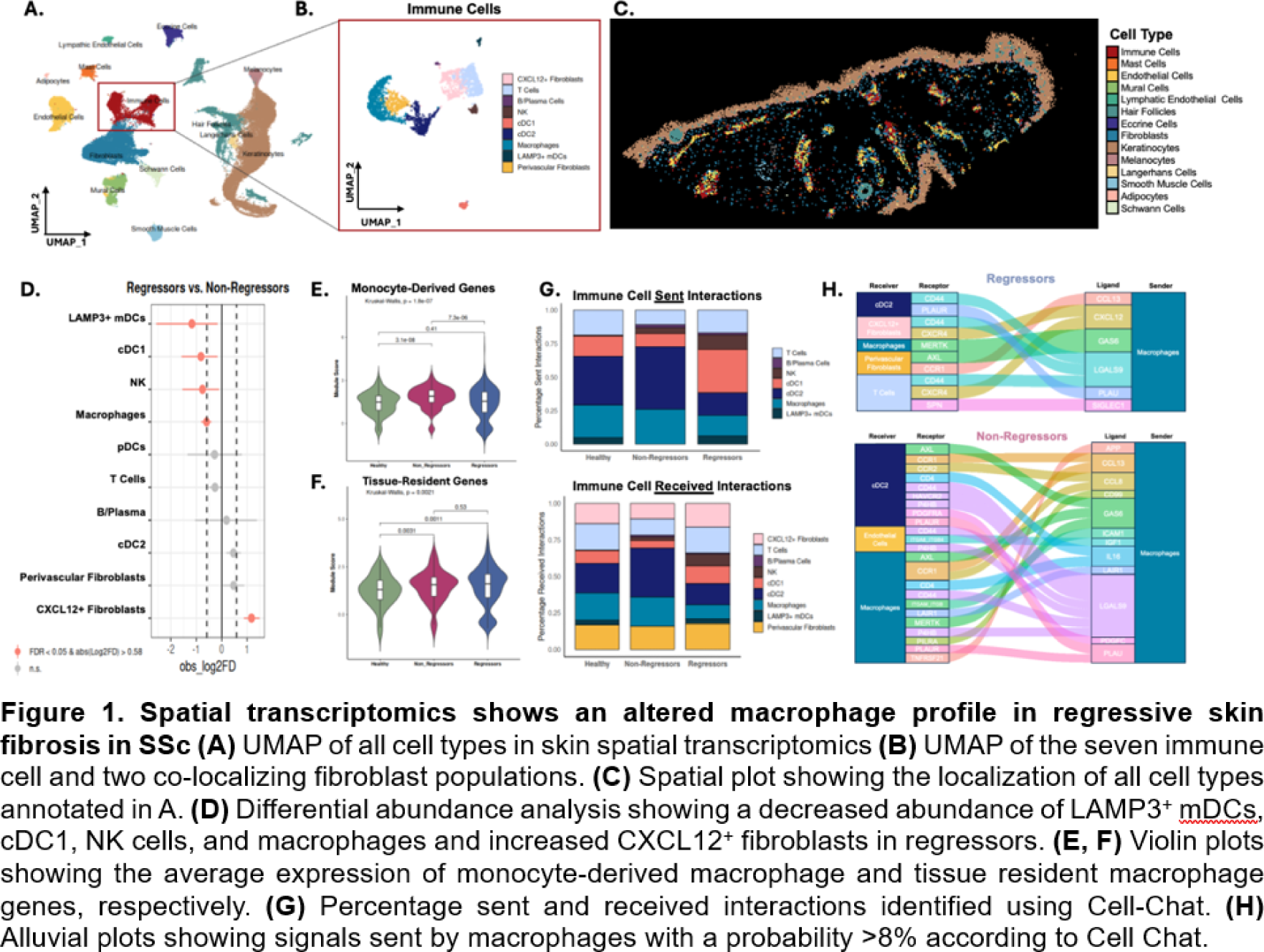
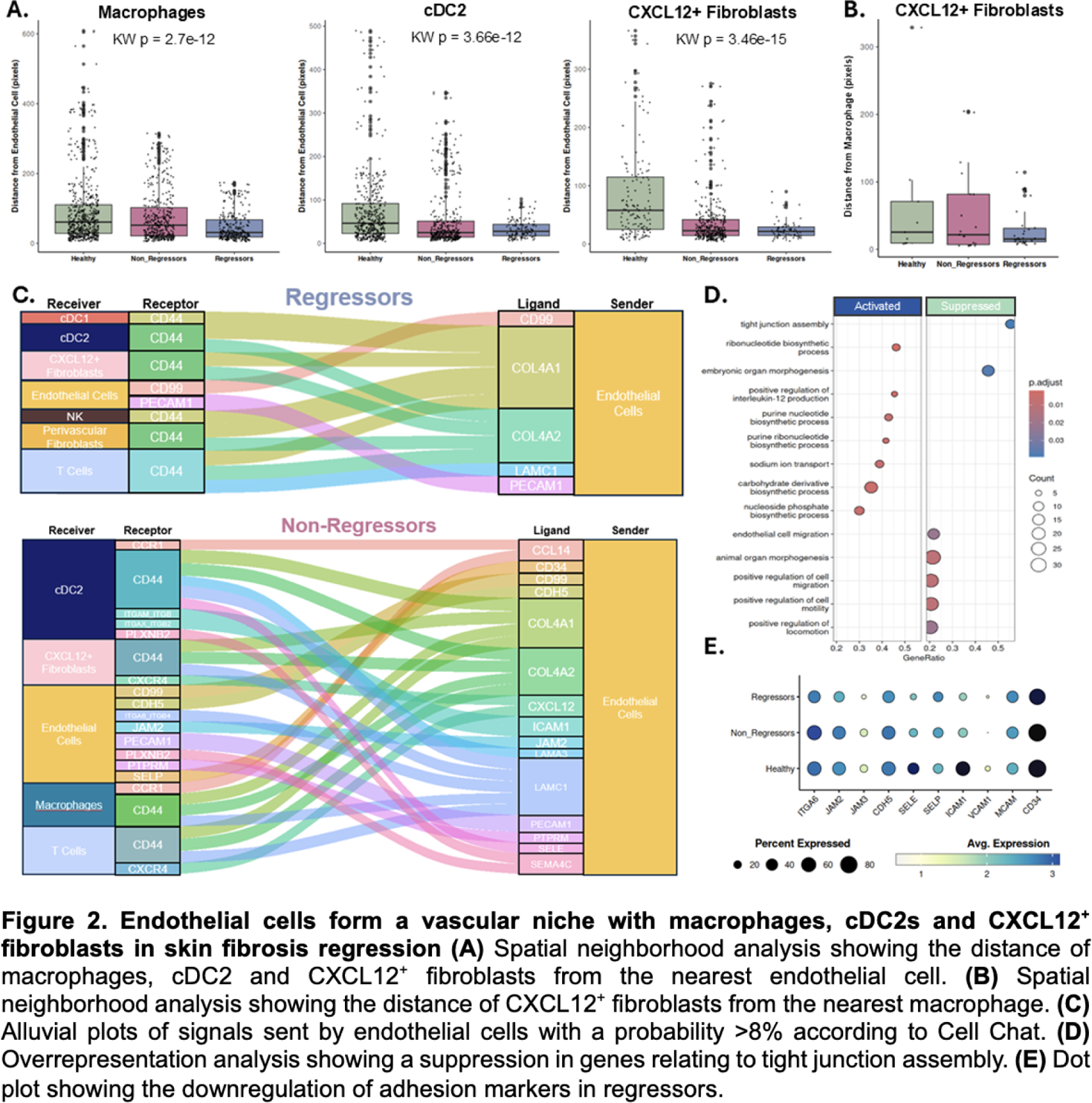
Results: Olink proteomics analysis identified 124 differentially abundant proteins between skin regressors (n=25) and non-regressors (n=19). Pathway enrichment analysis on differentially abundant proteins identified “cellular extravasation” and angiogenesis among the top enriched downregulated processes in regressors. Eighty-one percent of differentially abundant proteins in these processes were functionally linked to monocyte extravasation or modulation of endothelial cells. To assess if this signature was also present in the circulating immune cells, we used our PBMC scRNA-seq (n=10, 5 regressors, 5 progressors). This revealed five distinct clusters of monocytes (Mo1-Mo5). To identify extravasation-primed monocyte subpopulations the “Positive Regulation of Extravasation” gene set was used and found to be increased in the Mo1 (S100A8/9/12 + SELL + CD14 + , classical inflammatory) and Mo2 (HLA-DRA + CD74 + CIITA + CD14 + , antigen-presenting) subpopulations. Both the Mo1 and Mo2 subpopulations showed a suppression in genes relating to cell adhesion and motility in the regressors. Pathway enrichment analysis of the Mo1 and Mo2 populations demonstrated a downregulation in the interferon gamma (IFN-γ), interferon alpha (IFN-α), and TNF-α pathways in the regressors as compared to non-regressors. To assess whether this signature was maintained on tissue level, we used spatial transcriptomics to profile SSc skin biopsies (n = 12, 3 regressors, 5 non-regressors, 4 age/sex-matched healthy controls). 68,381 cells were retained after quality control, representing 14 major cell populations in the skin ( Figure 1A, C ). We identified seven immune cell and two fibroblast populations ( Figure 1B ). A reduced proportion of macrophages, conventional dendritic cells (cDC)1, LAMP3 + migratory DCs and natural killer cells was identified in the regressors, accompanied by an enrichment in CXCL12/SDF1 + fibroblasts ( Figure 1C ). Skin macrophages in regressors expressed significantly fewer monocyte-derived markers ( CD14, CCR2, MRC1 ) whereas tissue-resident macrophage signatures were unchanged, indicating that regressors specifically have fewer monocyte-derived inflammatory macrophages ( Figure 1E, F ). In regressors, cell-cell communication analysis showed a reduction in both sent and received interactions in the macrophages and cDC2s as compared to non-regressors ( Figure 1G ). Furthermore, regressors’ macrophages sent fewer signals but to a greater number of receiving immune and stromal populations. Conversely, in the non-regressors, the predicted interactions were enriched for chemotactic, inflammatory, and adhesion-related ligands ( CCL13, CCL8, IL16, ICAM1, PLAU ) (Figure 1H ). Overrepresentation analysis of endothelial cells in regressors showed marked suppression of adhesion molecules and tight junction–associated genes, including JAM3 and ACVRL1 , confirming our proteomics analysis (Figure 2D, E ). Non-regressors instead demonstrated robust endothelial outgoing signaling of chemotactic and adhesion-related signals ( CXCL12, CCL14, ICAM1, PECAM1, SELE ), reflecting an activated, immune-recruiting endothelium (Figure 2C ). Macrophages, cDC2s, and CXCL12 + fibroblasts were located perivascularly in regressors but not in non-regressors as identified by the distance to the nearest endothelial cells (Figure 2A ). When examining the distance from the macrophages, CXCL12 + fibroblasts were significantly closer to macrophages in regressors, suggesting a macrophage–fibroblast–endothelial niche (Figure 2B ).
Conclusions: Combining serum proteomics, PBMC scRNA-seq, and spatial transcriptomics, our data show that skin fibrosis regression is characterized by a systemic reduction in monocyte extravasation and a dampened inflammatory profile observed across biological compartments. In the skin, this signature is mirrored by a distinct vascular–immune–stromal niche in regressors characterized by altered immune cell localization and endothelial cell function.
REFERENCES: NIL.
Acknowledgments: NIL.
Disclosure of Interests: Astrid Hofman: None declared, Aleksandra Khmelevskaia: None declared, Pietro Bearzi: None declared, Cosimo Bruni Consultant for Boehringer Ingelheim and Glaxo Smith Klein, congress support from Boehringer Ingelheim, Laura Much: None declared, Maryam Asadikorayem: None declared, Blaz Burja consultant of Ely Lilly, Lumeng Li: None declared, Kristina Buerki: None declared, Muriel Elhai Boehringer Ingelheim, Vontobel Stiftung, Pfizer, Novartis Foundation for Bio-Medical Research, Iten Kohaut foundation, Kurt und Senta Herrmann foundation, Foundation for research in Rheumatology (FOREUM), University Zurich, Walter and Gertrud Siegenthaler Foundation. Congress Support from Janssen, Carina Mihai: None declared, Rucsandra Dobrota Actelion, Boehringer-Ingelheim, Actelion, Boehringer-Ingelheim, Iten-Kohaut, Walter und Gertrud Siegenthaler Fellowship, the LOOP Zurich, Pfizer, Actelion. Congress/Workshop Participation: Amgen, Otsuka, Sinziana Muraru Congress participation support AstraZeneca, Philip Stauffer: None declared, Anna-Maria Hoffmann-Vold Boehringer Ingelheim, Janssen, Medscape, Merck Sharp & Dohme, Novartis, Roche, AbbVie, Avalyn, Astra Zeneca, Boehringer Ingelheim, Bristol Myers Squibb, Calluna Pharma, Genentech, Janssen, Medscape, Merck Sharp & Dohme, Pliant, Roche, Werfen, Astra Zeneca, Boehringer Ingelheim, Janssen, Mike O. Becker Amgen, Vifor, Grant/research support from: Foundation for research in Rheumatology (FOREUM), EMDO Foundation, Novartis foundation for medical-biological research, Innovative Medicines Initiative (IMI)., GSK, Amgen, Novartis, Vifor. Congress support from Vifor., Elena Pachera: None declared, Oliver Distler Boehringer Ingelheim,: 4P-Pharma, Abbvie, Acepodia, Aera, AnaMar, Anaveon, Argenx, AstraZeneca, Avalyn, Boehringer Ingelheim, BMS, Calluna, Cantargia, CSL Behring, EMD Serono, Galderma, Galapagos, Gossamer, Hemetron, Innovaderm, Kali, Lilly, Mediar, MSD Merck, Nkarta, Novartis, Oorja Bio, Orion, Pliant, Prometheus, Quell, Scleroderma Research Foundation, Skyhawk, Tandem, Topadur, UCB and Umlaut.bio., Kymera, Mitsubishi Tanabe, UCB.