fetching data ... 
Background: The Sjögren’s Team for Accelerating Medicines Partnership (STAMP) aims to delineate the molecular mechanisms that drive Sjögren’s disease (SjD) pathogenesis. SjD is a heterogeneous autoimmune disease with systemic and organ-specific clinical and molecular manifestations, including serum anti-Ro autoantibody positivity and lymphocytic infiltration and destruction of exocrine glands. SjD lacks an established definition of early disease, further complicating clinical diagnosis and early intervention. Approximately 60-70% of SjD cases are serum anti-Ro/SSA autoantibody positive (Ro+), which correlates with increased systemic involvement, salivary gland lymphocytic infiltrations, and germinal center-like formation. Ro-negativity (Ro-) tends towards more organ-specific disease. To study early disease mechanisms, STAMP defined early SjD using these inclusion criteria: dryness symptoms onset £10 years earlier and preservation of some salivary gland tissue with minimal fibrosis, atrophy, and fatty infiltration; cases previously treated with disease-modifying antirheumatic drugs (DMARDs) or biologics or corticosteroid within 3 months of visit, and diagnosis of concomitant connective tissue disease were excluded.
Objectives: Leverage single-cell transcriptomics to define the molecular characteristics of early SjD subphenotypes in the salivary gland.
Methods: IRB approval obtained. Labial salivary gland (LSG) biopsies from 16 Ro+ SjD, 6 Ro- SjD, and 6 healthy controls (HCs) were subphenotyped by focus score (FS) – a quantitative measure of lymphocytic infiltration (Table 1). 10X Genomics scFlex was performed on single cells dissociated and captured from a 25mm section of formalin-fixed paraffin embedded (FFPE) LSG tissues. Sequencing data were analyzed (CellRanger), ambient RNAs corrected (DecontX), and doublets detected (scDblfinder). In total, 793,883 cells were captured. After normalization, cells were clustered and annotated using a 5-tier approach. Major epithelial, mesenchymal and immune cell clusters were annotated in Tier 1. Subsequent tiers annotated cluster subsets with final manual annotation of 56 cell types and states in Tier 5. After final QC, 305,843 total cells were annotated. Differential cell composition across subphenotypes were analyzed, excluding samples with <2000 cells. Differential gene expression (DEG) was performed on pseudo-bulk cell types (DESeq2), filtering for ≥10 cells in each cell type per subject and genes with ≥10 counts per sample. Pathway Analysis (IPA) was performed using DEGs.
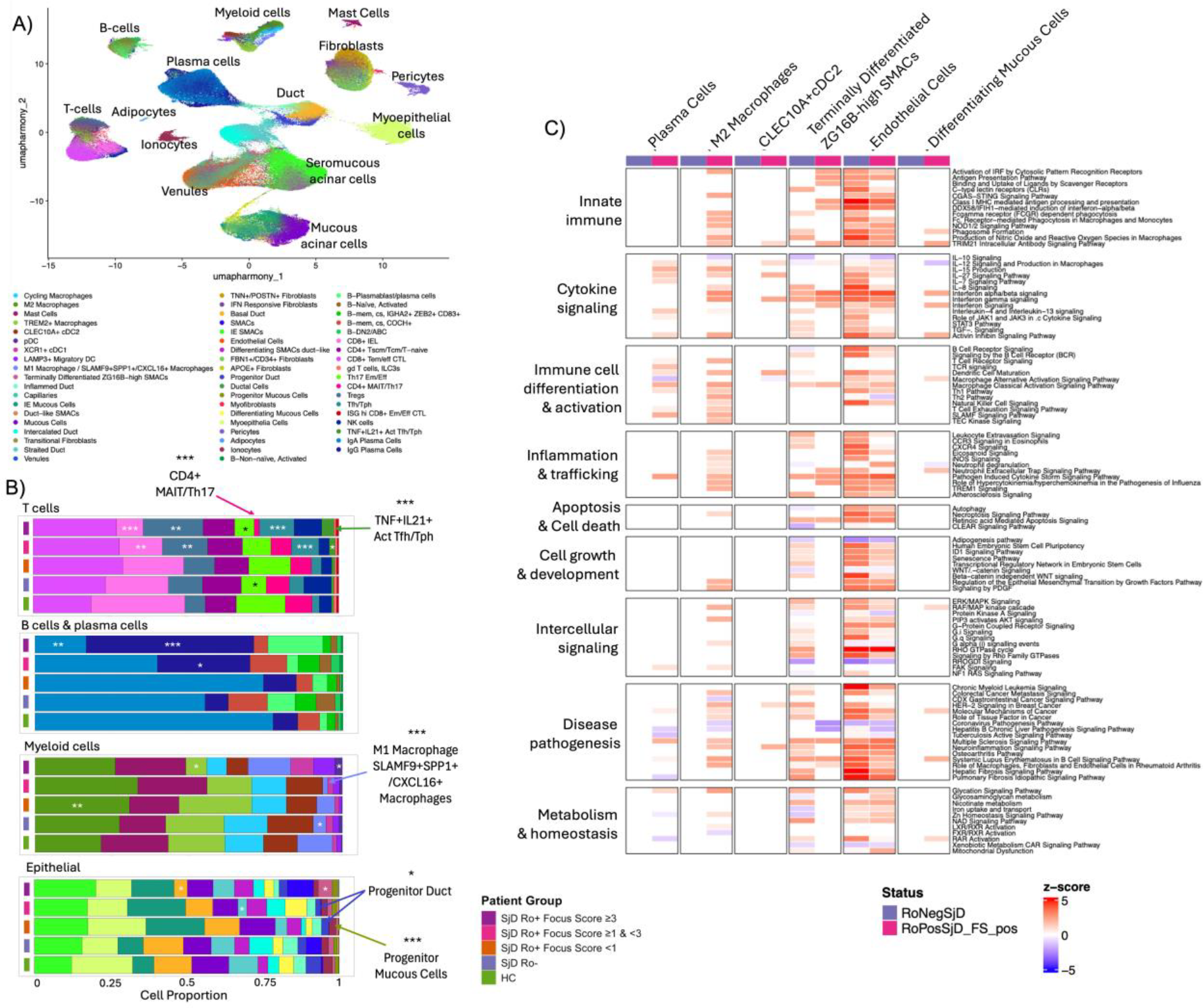
Results: scFFPE-seq analysis of LSG captured 56 distinct cell types (Figure 1A) and revealed subphenotype-specific cell proportion differences (Figure 1B). In the T cell population, the proportion of T helper 17 effector memory/effector cells (Th17 Em/Eff) were reduced in the Ro- SjD and Ro+ SjD FS>3 groups compared to HC. CD8+ intraepithelial lymphocytes (IELs) were also reduced in Ro+ SjD FS>3 compared to HC. In contrast, TNF+ IL21+ activated T follicular helper (Tfh)/T peripheral helper (Tph) cells, Tfh/Tph cells, and regulatory T cells (Tregs) were proportionally increased in Ro+ SjD with higher FS. Similar increases were also observed in Ro+ SjD plasma cells in samples with higher FSs. In the myeloid lineage, Ro- SjD and Ro+ SjD FS>1 showed lower SLAMF9+SSP1+/CXCL16+ macrophages but trended toward higher M2 macrophages compared to HCs; however, only Ro+ SjD FS<1 reached significance. Interestingly, the Ro+ SjD FS>3 showed significantly fewer TREM2+ macrophages compared to HC. In the epithelial cluster, Ro+ SjD FS<1 and FS <3, but not Ro- SjD, showed significant reductions in progenitor duct and mucous cells. Pathway analysis showed increased cytokine signaling and IRF pathways in plasma cells, M2 macrophages, CLEC10A+ pDC2, terminally differentiated ZG16B high seromucous acinar cells (SMACs), and endothelial cells from SjD Ro+ FS+ (Figure 1c). Immune related pathways were similarly upregulated in endothelial cells and terminally differentiated ZG16B high SMACs from SjD Ro- (Figure 1C). No significant changes in shown pathways and cell types were observed in SjD Ro+ FS- compared to HC.
Conclusions: This study is the first of a multi-phase effort to expand cohort size, incorporate longitudinal sampling, and enable more robust testing of disease trajectories and mechanistic hypotheses during SjD pathogenesis. It established the scFFPE-seq workflow and resolved 56 cell states in LSG, focusing on early SjD. Ongoing cell type-specific analyses reveal complex immune and epithelial cell states in LSG, such as increased helper and regulatory T cell populations associated with germinal center activity and chronic immune activation. Reduced proportions of mucous progenitor and ductal cells in Ro+ SjD FS- compared to HC may reflect early impairment of glandular regenerative capacity. One SjD Ro+ case with FS=12 showed elevated plasmablast and plasma cells, indicative of severe inflammation and potential shift to autoreactive antibody production. Future studies will include additional high FS cases to allow for detailed study of this cell population in the context of SjD.
Sjögren’s disease (SjD) cases and healthy controls (HCs) subphenotyped by anti-Ro/SSA positivity and focus score (FS) of the labial salivary gland.
| SjD Ro+ (n=16) | SjD R- (n=6) | HC (n=6) | ||||
|---|---|---|---|---|---|---|
| Criteria | FS<1 | 3>FS≥1 | FS≥3 | FS>1 | FS<0.3 | FS≥0.3 |
| Samples (n) | 4 | 7 | 5 | 6 | 3 | 3 |
| FS Median [min-max] | 0.1 [0-0.75] | 1.8 [1.25-2.25] | 5.5 [3.2-12] | 1.675 [1.05-3.6] | 0 [0-0.25] | 0.8 [0.55-0.85] |
scFFPE-seq of Minor Salivary Gland (MSG). (A) Cell type identification using UMAP and manual annotation. (B) Composition analysis for T cells, B cells & plasma cells, myeloid cells and epithelial cells. Stacked bar plots show cell proportions across the indicated groups. Differences in cell population proportions were tested using propeller compared with HC: * p < 0.05, ** p < 0.01, *** p < 0.001. (C) Pathway analysis for the indicated LSG cell types, grouped by SjD Ro-, SjD Ro+ FS-, and SjD Ro+ FS+.
REFERENCES: NIL.
Acknowledgments: NIL.
Disclosure of Interests: Marcin Radziszewski: None declared, James Li: None declared, Songyuan Yao: None declared, Cherilyn Pritchett Frazee: None declared, Mandi M Wiley: None declared, Kandice L Tessneer: None declared, Matthew Caleb Marlin Received consulting fees from Standard Biotools., Miles Smith: None declared, Kevin Wei: None declared, Soumya Raychaudhuri: None declared, Michael Brenner: None declared, Lam Tsoi: None declared, Johann E Gudjonsson: None declared, Nir Hacohen: None declared, Umesh Deshmukh Grant/research support from Johnson and Johnson Innovative Medicine (formerly Janssen; ended 12/31/2023), Susan Kovats: None declared, Judith A. James Received consulting fees from GSK, Received research support from Progentec Biosciences, R Hal Scofield Advisory Board Member for Janssen Pharmaceuticals, IQVIA, Received consulting fees from Johnson and Johnson Innovative Medicine (formerly Janssen) and Merk Pharmaceuticals, Astrid Rasmussen Current affiliation: Boston Children’s Hospital, Received consulting fees from Immunovant Clinical Outcomes Solutions, Stephen Shiboski: None declared, Bhuwan Khatri: None declared, Alan Baer Received consulting fees from Bristol Myers Squibb (BMS), A. Darise Farris Grant/research support from Johnson and Johnson Innovative Medicine (formerly Janssen; ended 12/31/2023), Caroline Shiboski Received consulting fees from BMS, Joel M Guthridge: None declared, Blake M Warner Active research agreements with Astellas Bio, Pfizer, Christopher J Lessard Consulting fees from Johnson and Johnson Innovative Medicine Research Alliance Sjögren’s Disease Advisory Board, Received research support from Johnson and Johnson Innovative Medicine (formerly Janssen; ended 12/31/2023).