fetching data ... 
Background: Systemic lupus erythematosus (SLE) is a heterogeneous autoimmune disease characterized by loss of self-tolerance, systemic inflammation, and multi-organ involvement. Sustained activation of the type I interferon (IFN-I) pathway, particularly IFN-α, is a central pathogenic mechanism, promoting B cell maturation, plasmablast differentiation, antigen presentation, and inflammatory T cell polarization [1]. Anifrolumab, a monoclonal antibody targeting the IFNAR1 receptor, blocks IFN-I signaling and has demonstrated efficacy in reducing disease activity, flares, and corticosteroid use in clinical trials [2-4]. Bulk transcriptomic analyses show downregulation of interferon-stimulated genes and restoration of regulatory immune signatures [4], but cell-specific therapeutic effects remain incompletely understood. Single-cell and spatial transcriptomic approaches provide high-resolution insight into immune responses, both in circulation and affected tissues, highlighting the importance of dissecting systemic and tissue-specific mechanisms of IFN-I blockade.
Objectives: To investigate how Anifrolumab modulates immune cell composition and interferon signaling in PBMCs and skin of SLE patients, integrating systemic and tissue-specific single-cell and spatial transcriptomic analyses.
Methods: Peripheral blood mononuclear cells (PBMCs) were collected from 15 SLE patients treated with Anifrolumab at three time points: pre-treatment (flare), 3–6 months post-treatment, and 9–12 months post-treatment. Five healthy donors were included as controls. Skin biopsies were obtained from three SLE patients at flare and at 9–12 months post-treatment, including paired non-lesional skin, along with two healthy controls. Single-cell transcriptomic profiling of PBMCs was performed using CITE-seq (BD Rhapsody), and spatial transcriptomic profiling of skin samples was performed using CosMx Spatial Transcriptomics (1000-plex). Immune cell composition, transcriptomic changes, and interferon signaling pathways were analyzed longitudinally in both blood and tissue.
Results: At flare, SLE patients exhibited significantly higher frequencies of naïve CD4 and CD8 T cells compared with healthy donors (fold change 1.8 and 2.7, respectively). Following treatment, these naïve populations decreased significantly (fold change 1.4), while CD4 effector memory T cells (TEM) expanded (fold change 2.1). Additionally, after three months of treatment, classical monocytes and CD56^medium natural killer cells increased (fold change 1.7 and 1.4), reflecting early remodeling of both innate and adaptive compartments.
Transcriptomic profiling revealed broader changes at the early treatment time point, particularly in TEM and naïve T cells, characterized by downregulation of pathways related to translation, viral transcription and replication, nonsense-mediated decay, ribosomal assembly, and interferon-regulated antiviral responses. At the latest post-treatment time point, additional transcriptomic alterations were observed in naïve and switched memory B cells, naïve T cells, and myeloid populations. In B cells, these changes included downregulation of three major interferon-related pathways (interferon alpha/beta signaling, general interferon signaling, and antiviral mechanisms mediated by interferon-stimulated genes) accompanied by activation of mRNA translation pathways. Naïve T cells primarily exhibited suppression of interferon signaling, whereas myeloid cells preferentially downregulated interferon gamma–associated pathways.
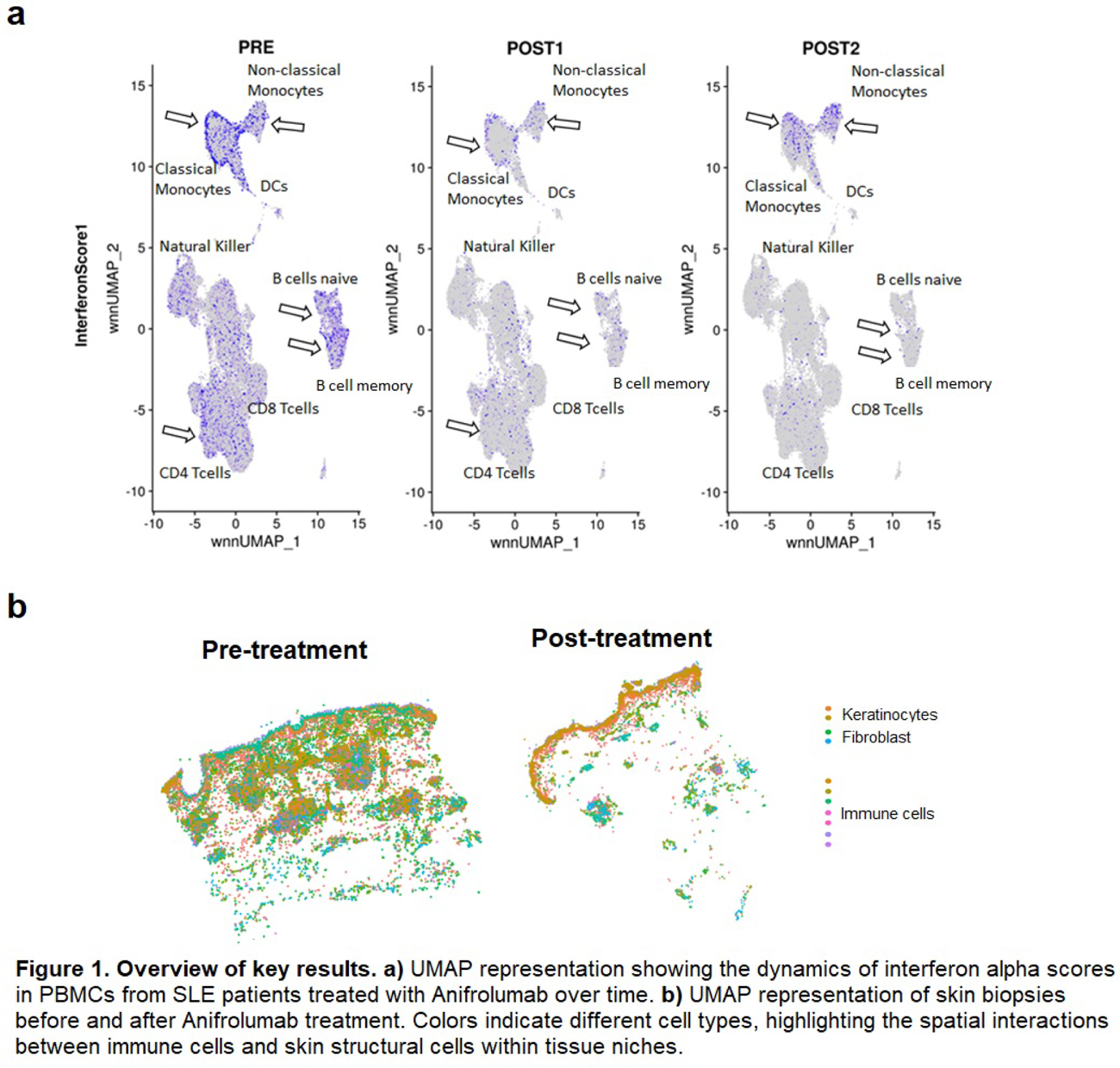
Longitudinal analysis of interferon scores showed that interferon gamma remained stable, while interferon alpha and kappa scores varied significantly across classical, non-classical, and intermediate monocytes, B cells, CD4 T cells and dendritic cells. By 12 months post-treatment, interferon scores were restored in all monocyte subsets (Figure 1a).
Analysis of skin biopsies revealed increased abundance of myeloid dendritic cells, CD8-NKT cells, and macrophages in pre-lesional skin compared with non-lesional or healthy control skin. Myeloid dendritic cells and macrophages were the main producers of interferon alpha, while naïve and memory B cells also contributed; all these populations showed significant reductions after treatment. Keratinocytes were the primary source of interferon kappa, which was similarly decreased post-treatment. Spatial analysis revealed close interactions between skin-resident immune cells and epidermal or stromal cells, indicating that interferon signaling was locally reduced within specific tissue niches (Figure 1b).
Conclusions: Anifrolumab treatment in SLE patients induces coordinated modulation of the immune system both systemically and in tissue. Early effects primarily target naïve and effector T cells, downregulating translation- and interferon-related pathways, while later effects extend to B cells and myeloid populations. Interferon alpha/kappa scores and interferon-producing cell populations in PBMCs and skin normalize following treatment, highlighting the therapy’s ability to suppress pathogenic interferon signaling and restore immune homeostasis. These findings provide mechanistic insight into Anifrolumab’s targeted action in SLE and demonstrate its capacity to modulate both systemic and tissue-specific interferon responses.
REFERENCES: [1] Wahren-Herlenius M, et al. Immunopathogenic mechanisms of systemic autoimmune disease. Lancet 2023 ;382:819-31.
[2] Morand EF, et al. Trial of anifrolumab in active systemic lupus erythematosus. N Engl J Med. 2020 ;382:211-221.
[3] Chatham WW, et al. Long-term safety and efficacy of anifrolumab in adults with systemic lupus erythematosus: results of a phase II open-label extension study. Arthritis Rheumatol 2021 ;73:816-825.
[4] Baker T, et al. Type I interferon blockade with anifrolumab in patients with systemic lupus erythematosus modulates key immunopathological pathways in gene expression and proteomic analysis of two phase 3 trials. Ann Rheum Dis 2024 ;83:1018-1027.
Acknowledgments: NIL.
Disclosure of Interests: None declared.