fetching data ... 
Background: Systemic lupus erythematosus (SLE) is characterized by impaired regulatory T cell (Treg) function. Low-dose interleukin-2 (LD-IL-2) can restore immune tolerance by selectively activating and expanding Tregs via their high-affinity receptors. While our previous studies established that LD-IL-2 at 0.5 million International Units [MIU]/day improves Treg homeostasis and alleviates clinical symptoms in SLE, the precise cellular functional dynamics and intercellular regulatory networks remain incompletely understood.
Objectives: Leveraging single-cell RNA sequencing (scRNA-seq), this study sought to delineate the longitudinal remodeling of the peripheral immune microenvironment in SLE patients responding to LD-IL-2 therapy.
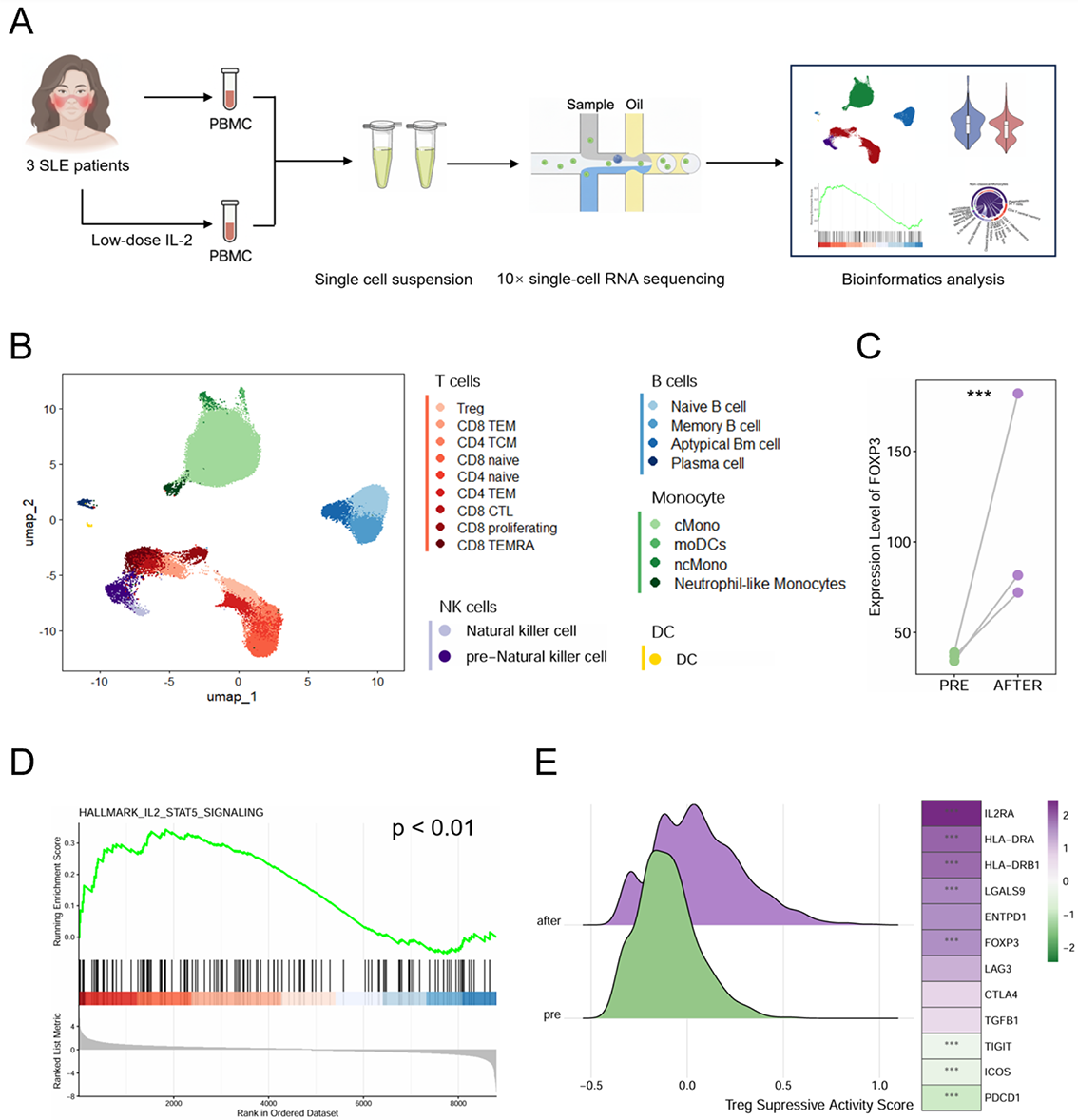
Methods: Three newly diagnosed SLE patients received low-dose IL-2 monotherapy (0.5 MIU/day for five consecutive days) and exhibited significant clinical improvement. Peripheral blood mononuclear cells (PBMCs) were collected longitudinally at baseline (pre-treatment) and post-treatment for scRNA-seq ( Figure 1A ). Data were processed using CellRanger and Seurat. Functional characteristics were assessed using the AddModuleScore function and GSEA via clusterProfiler, while intercellular signaling was inferred using CellChat.
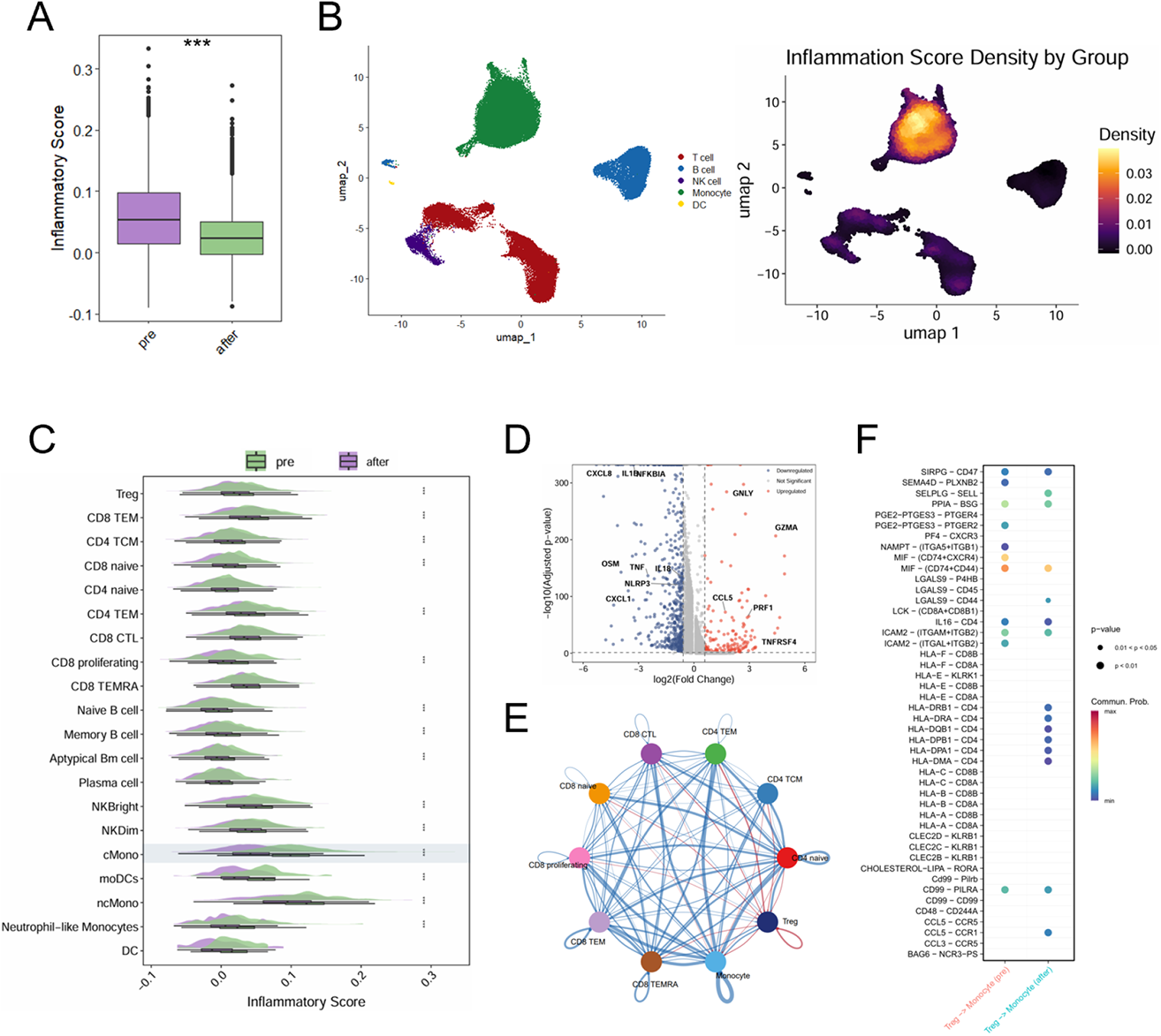
Results: Following rigorous quality control, analysis of 51,254 single-cell transcriptomes were analyzed and classified into 20 subsets across 5 major immune lineages ( Figure 1B ). Post-treatment Tregs exhibited a reinforced suppressive phenotype characterized by enhanced FOXP3 expression and enriched IL-2-STAT5 signaling. This functional enhancement was further supported by upregulation of IL2RA and HLA-DRB1, indicative of an activated, immunoregulatory state ( Figure 1C-E ). Although overall global inflammatory scores decreased after therapy, the most profound attenuation occurred within the monocyte compartment ( Figure 2A-C ), characterized by the widespread downregulation of key inflammatory mediators including TNF, IL1B, and CXCL8 ( Figure 2D ). Monocytes expressed negligible IL2RA, suggesting that this shift is not a direct consequence of IL-2 signaling. Instead, cell-communication analysis revealed substantial rewiring of Treg-monocyte interactions ( Figure 2E ). Specifically, we observed the emergence of the immunosuppressive LGALS9 signaling, indicative of enhanced Treg inhibitory function. Concurrently, the pro-inflammatory MIF signaling pathway was abrogated, thereby fostering an immune-tolerant microenvironment ( Figure 2F ). These analysis of interactions between Treg and Monocyte suggested anti-inflammatory reprogramming of monocyte by Treg cells after LD-IL-2 treatment via LGALS9 and MIF signaling.
Conclusions: Our single-cell transcriptomic landscape elucidates that LD-IL-2 restores immune tolerance by qualitatively potentiating Treg function via the IL-2-STAT5 axis and indirectly reprograms monocytes toward an anti-inflammatory state through remodeled Treg-monocyte crosstalk. These findings highlight the capacity of LD-IL-2 to coordinately modulate adaptive and innate immunity in SLE, providing a mechanistic foundation for future therapies targeting intercellular communication networks.
scRNA -seq analysis and identification of PBMC cell subpopulations in SLE patients undergoing LD-IL-2 treatment
A. Schematic overview of the study design. B. Uniform Manifold Approximation and Projection (UMAP) visualization of the integrated immune landscape. C. Paired comparison of FOXP3 expression levels in Tregs between pre- and post-treatment. D. Gene Set Enrichment Analysis (GSEA) plot demonstrating the significant enrichment of the “HALLMARK_IL2_STAT5_SIGNALING” pathway in post-treatment Tregs compared to pre-treatment. E. Functional characterization of Tregs. Left: Density plot showing the distribution of Treg suppressive activity scores. Right: Heatmap displaying the scaled expression of key functional and suppressive markers. *** p<0.001.
LD-IL-2 treatment induces anti-inflammatory gene expression signature of monocytes
A. Box plot comparing the global inflammatory scores of PBMCs between pre- and post-treatment groups. B. UMAP visualization of the immune landscape. Left: Color-coded by major cell lineages. Right: Density plot of inflammatory scores. C. Split violin plots showing the distribution of inflammatory scores across cell subsets pre- and post-treatment. D. Volcano plot displaying differentially expressed genes in classical monocytes (Post vs. Pre). E. Circle plot illustrating the aggregated cell-cell communication network among major immune cell subsets. F. Dot plot comparing significant ligand-receptor interactions from Tregs to Monocytes inferred. *** p<0.001.
REFERENCES: NIL.
Acknowledgments: NIL.
Disclosure of Interests: None declared.