fetching data ... 
Background: The pivotal role of CD40-CD40L interactions in systemic lupus erythematosus (SLE) pathogenesis stems from the orchestration of a range of immune and inflammatory responses involving T cells, B cells, and other antigen-presenting cells, which result in the production of pro-inflammatory cytokines and autoantibodies [1]. Dapirolizumab pegol (DZP) is a novel, polyethylene glycol (PEG)-conjugated antigen-binding fragment (Fab’), lacking an Fc domain. DZP binds CD40L, blocking CD40-CD40L interactions and CD40 activation, and has broad modulatory effects on SLE immunopathology [2, 3, 4]. DZP demonstrated significant efficacy and was generally well tolerated in patients with SLE in the phase 3 PHOENYCS GO trial (NCT04294667) [5], and is under investigation in a further phase 3 clinical trial (PHOENYCS FLY; NCT04976322). Post hoc analyses of the phase 2b RISE trial (NCT02804763) of DZP in SLE demonstrated that DZP suppresses the expression of genes involved in B and T cell activation, immunoglobulin production, and pro-inflammatory cytokines in patients with active SLE [3, 4].
Objectives: To investigate the effect of DZP on in vitro T and B cell interactions, and cytokine and immunoglobulin production, relevant to SLE pathogenesis.
Methods: Antibody induction T follicular helper (Tfh)-B cell co-culture experiments were performed using peripheral blood mononuclear cells (PBMCs) from healthy volunteers (N=6) isolated by Ficoll-Paque. Naïve T cells, Tfh cells, and naïve B cells were further purified by flow cytometric sorting. Naïve T cells or Tfh cells were then incubated with naïve B cells at a 1:1 ratio in the presence of 1 μg/mL Staphylococcal enterotoxin B (SEB) for 14 days with DZP or A33 Fab’-PEG isotype control (15 μg/mL). Total or SEB-specific IgG and IgM were measured by MesoScale Discovery (MSD) and enzyme-linked immunosorbent assay (ELISA), respectively. Intracellular IFN-γ production experiments were performed using whole blood samples from patients with SLE (N=12). Red blood cells were lysed and cultured for 18 hours with 1 μg/mL SEB and DZP or A33 Fab’-PEG isotype control (100 μg/mL) in the presence of Brefeldin A and Monensin to prevent cytokine secretion. Intracellular cytokine staining of CD4+ and CD8+ T cells was performed 18 hours after SEB antigen stimulation. PBMC cytokine production experiments were performed using whole PBMCs isolated by Ficoll-Paque from healthy volunteers (N=10) or patients with SLE (N=15). PBMCs were cultured in the presence of 1 μg/mL SEB for 5 days with DZP or A33 Fab’-PEG isotype control (100 μg/mL). Cytokine production, measured by LEGENDplex, was assessed after 5 days of culture with SEB. Data were analysed using a three-way ANOVA based on treatment, SEB-stimulation, and sample type. Comparisons between DZP and Fab’-PEG control were assessed for each combination of SEB-stimulation and sample type with t-tests using a pooled estimate of variance from the ANOVA.
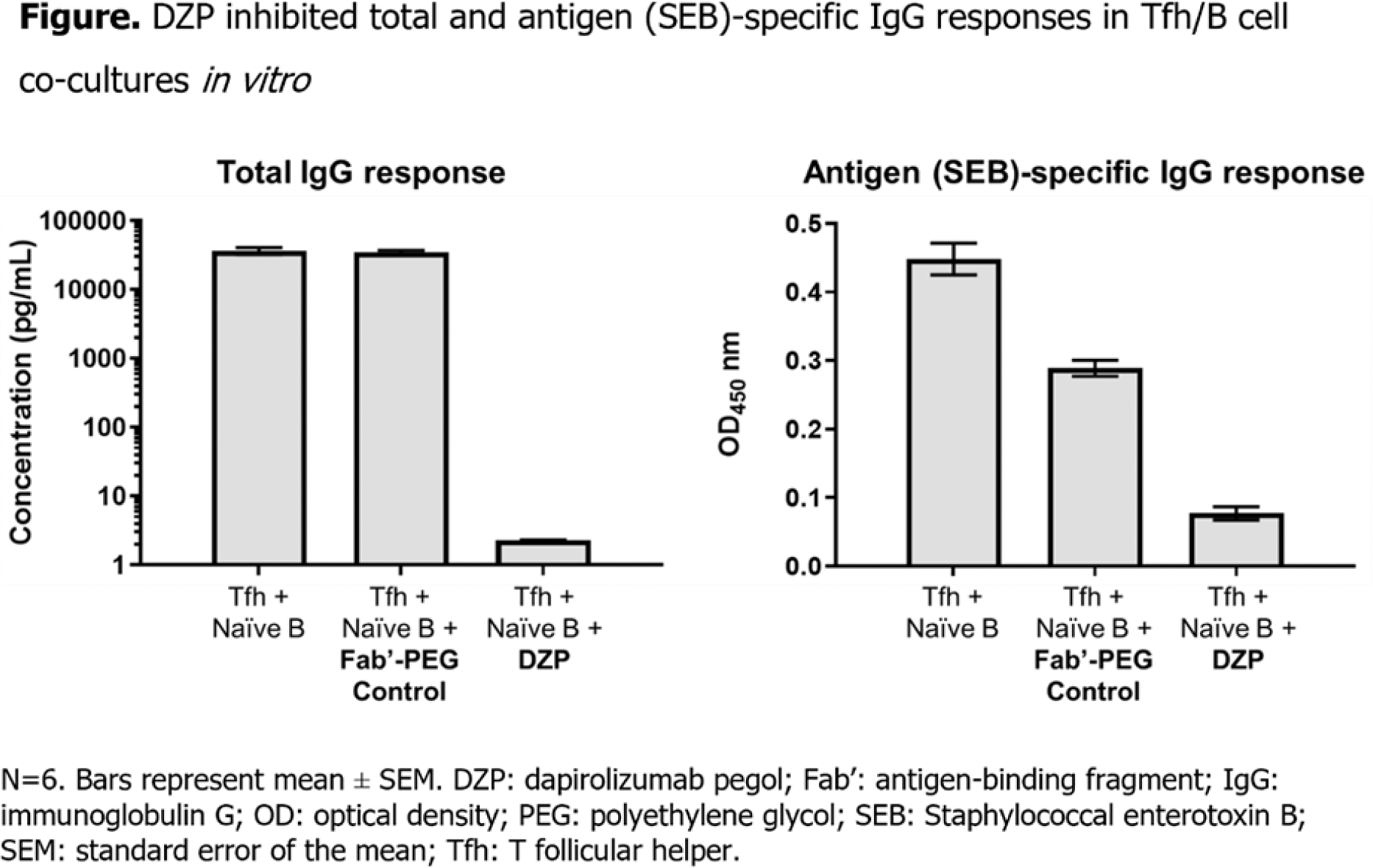
Results: Healthy volunteer Tfh cells were shown to drive increased naïve B cell antibody responses, increased IgG and IgA production, and class switching over naïve T cells in an in vitro co-culture assay. DZP inhibited production of both total and antigen (SEB)-specific IgG antibody responses relative to the isotype control (Figure 1). IFN-γ is a key pro-inflammatory cytokine involved in SLE pathogenesis and DZP was shown to reduce induction of IFNγ+ CD4+ and CD8+ T cells from patients with SLE after SEB stimulation relative to the isotype control. After culture of SEB-stimulated PBMCs isolated from both healthy volunteers and patients with SLE, DZP statistically significantly inhibited the release of a range of cytokines, including IL-1ß, IL-10, IL-12p40, and IL-6, relative to the isotype control (p<0.05).
Conclusion: DZP was shown to inhibit key T cell, B cell, and APC cell functions that drive SLE pathogenesis, leading to suppression of cytokines, and total and antigen-specific IgG production in vitro . These data support clinical and transcriptomic/proteomic findings from patients with SLE treated with DZP, where reductions of immunoglobulins and key pro-inflammatory cytokines have been observed [3, 4, 6, 7]. Overall, the data indicate that DZP modulates multiple pathways relevant to the pathogenesis of SLE.
REFERENCES: [1] Ramanujam M. Autoimmun Rev 2020;19:102668.
[2] Shock A. Arth Res Ther 2015;17:234.
[3] Cutcutache I. Arthritis Rheumatol 2023;75 (suppl 9).
[4] Powlesland A. Annals Rheum Dis 2024;83 (suppl 1):261.
[5] Clowse M. Arthritis Rheumtol 2024;76 (suppl 9).
[6] Furie R. Rheumatology (Oxford). 2021;60:5397–407.
[7] Chamberlain C. Ann Rheum Dis 2017;76:1837–44.
Acknowledgements: TR and ARK contributed equally to the conduct of the study. This study was funded by UCB and Biogen. Medical writing support provided by Costello Medical and funded by UCB and Biogen.
Disclosure of Interests: Tania Rowley Shareholder of UCB, employee of UCB, Adnan Khan Former employee of UCB, Laura McLaughlin Former employee of UCB, Hannah Cherry Shareholder of UCB, employee of UCB, Farnaz FALLAH-ARANI Shareholder of UCB, employee of UCB, John Ioannou Shareholder of UCB, employee of UCB, Ania Skowera Shareholder of UCB, employee of UCB, Debasish Pyne: None declared, Anthony Shock Shareholder of UCB, employee of UCB.
© The Authors 2025. This abstract is an open access article published in Annals of Rheumatic Diseases under the CC BY-NC-ND license (